

Bioactive Metabolites from Terrestrial and Marine Actinomycetes
by
 , , , and
, , , and
Chananan Ngamcharungchit
1,2,
Nutsuda Chaimusik
1,2,
Watanalai Panbangred
3,
Jirayut Euanorasetr
4,5,* and
Bungonsiri Intra
1,2,* 1
Department of Biotechnology, Faculty of Science, Mahidol University, Bangkok 10400, Thailand
2
Mahidol University and Osaka University Collaborative Research Center on Bioscience and Biotechnology, Bangkok 10400, Thailand
3
Research, Innovation and Partnerships Office, King Mongkut’s University of Technology Thonburi, Bangkok 10140, Thailand
4
Department of Microbiology, Faculty of Science, King Mongkut’s University of Technology Thonburi, Bangkok 10140, Thailand
5
Laboratory of Biotechnological Research for Energy and Bioactive Compounds, Department of Microbiology, Faculty of Science, King Mongkut’s University of Technology Thonburi, Khet Thung Khru, Bangkok 10140, Thailand
*
Authors to whom correspondence should be addressed.
Molecules 2023, 28(15), 5915; https://doi.org/10.3390/molecules28155915
Submission received: 16 June 2023
/
Revised: 27 July 2023
/
Accepted: 1 August 2023
/
Published: 6 August 2023
(This article belongs to the Special Issue Advances in Natural Products and Their Biological Activities)
Abstract
:Actinomycetes inhabit both terrestrial and marine ecosystems and are highly proficient in producing a wide range of natural products with diverse biological functions, including antitumor, immunosuppressive, antimicrobial, and antiviral activities. In this review, we delve into the life cycle, ecology, taxonomy, and classification of actinomycetes, as well as their varied bioactive metabolites recently discovered between 2015 and 2023. Additionally, we explore promising strategies to unveil and investigate new bioactive metabolites, encompassing genome mining, activation of silent genes through signal molecules, and co-cultivation approaches. By presenting this comprehensive and up-to-date review, we hope to offer a potential solution to uncover novel bioactive compounds with essential activities.
1. Introduction
Actinomycetes are generally recognised as filamentous Gram-positive bacteria in the order Actinomycetales [1]. Typical actinomycete colonies show distinct powdery characteristics containing filamentous mycelium-like fungi and spore-forming properties (e.g., genera Streptomyces, Microbispora, Streptosporangium, and Microbispora). The genus Streptomyces stands out as the most dominant and well-known actinomycetes. These are primarily aerobic bacteria with a notable high G + C content in their DNA, approximately ranging from 60% to 78% [1,2]. The Actinomycetales members show extensive diversity in their morphology, physiology, and metabolic capabilities. Rare or non-streptomycete actinomycetes are a group of actinomycete bacteria that are rarely isolated from the environment compared to dominant Streptomyces. Rare actinomycetes include Actinomadura, Actinoplanes, Actinokineospora, Actinosynema, Kineosporia, Planobispora, Nocardia, Thermomonospora, Saccharothrix, and Saccharopolyspora [3]. Rare actinomycetes have attracted attention due to their low frequency in isolation and the potential discovery of new natural bioactive compounds such as macrolide antibiotics [4,5].
Most actinomycetes are aerobic, saprophytic microorganisms with complex life cycles (Figure 1) except for unicellular Corynebacterium and Mycobacterium [6]. Actinomycetes have well-developed radial mycelium dividing into the substrate and aerial mycelium during their life cycle. Substrate mycelium is developed in the media to assimilate nutrients, and then aerial mycelium is developed afterwards. However, when actinomycetes grow in an impoverished environment, the hypha becomes coiled and develops a septum. After the septum develops, conidiospores form within the hyphae. Except for Streptomyces with its long-chain spores, other genera have distinguishable characteristics, such as Micromonospora (single non-motile spore), Microbispora (two spores in a chain), and Streptosporagium (bearing sporangium as spore vesicle) [1]. Spores will be released into the environment and germinate when favourable condition is achieved. In the germination step, the spore will protrude germ tubes then, germ tubes reach the vegetative growth stage, and the cycle is repeated [2,7].
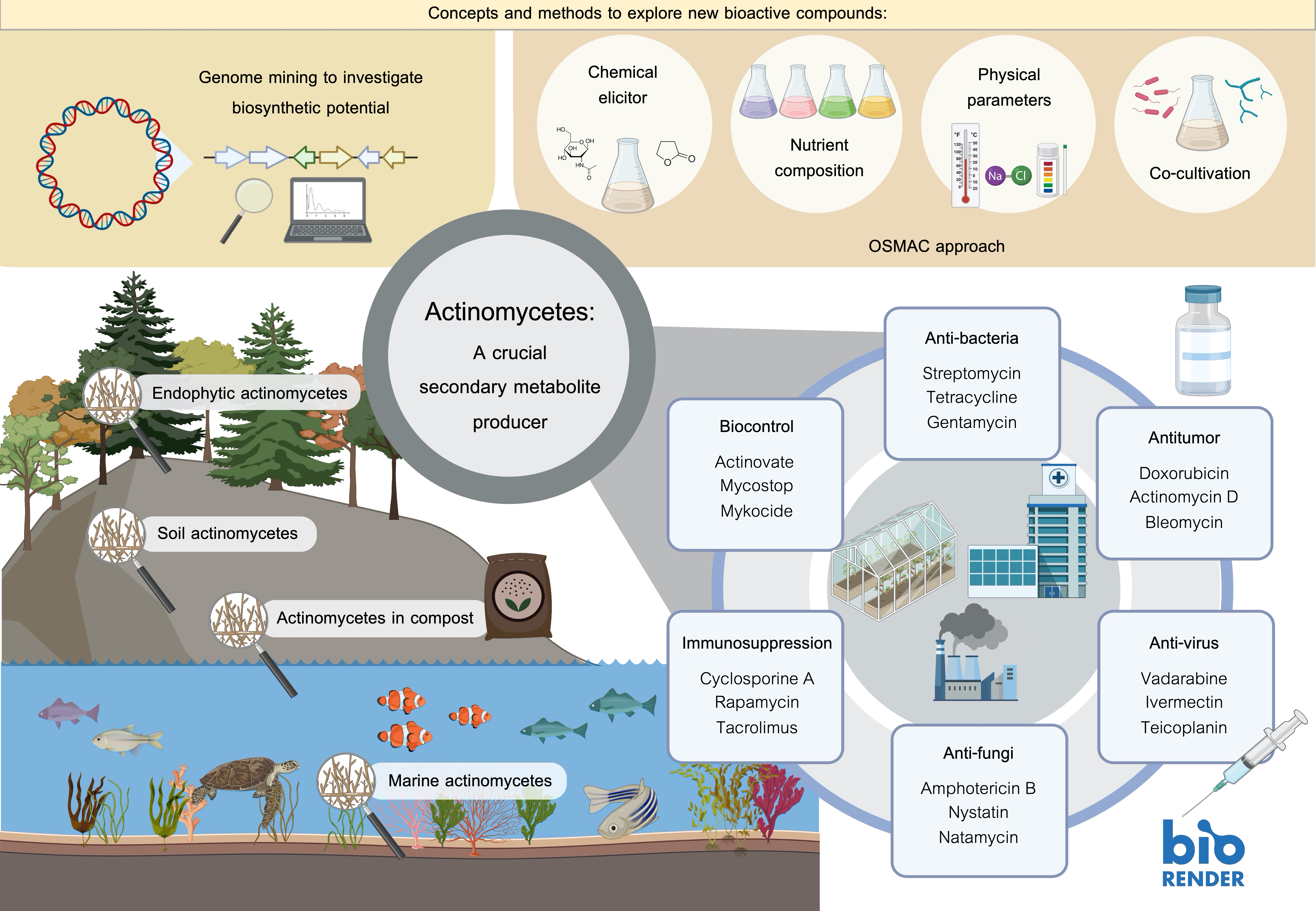
Actinomycetes play an important role in biotechnology and pharmacology because they can produce various secondary metabolites and other useful compounds such as antibiotics, antitumor agents, immunosuppressive agents, and nutritional materials. Therefore, their secondary metabolites and diversity make actinomycetes an important group of key compound producers. This review focuses on the recent bioactive metabolites derived from terrestrial and marine actinomycetes from the years 2016 to 2023. Additionally, the methodology for discovering new metabolites is also discussed.
2. Ecology of Actinomycetes
Actinomycetes are widely distributed in various ecosystems and habitats including soil and marine habitats, freshwater, animals, plants, insects, and fertilizer (Figure 2) [8,9]. They are free-living, saprophytes in an environment such as soil pore [8] or living as endophytes in plants [10]. The actinomycete members include inhabitants of soil or aquatic environments (e.g., Streptomyces, Micromonospora, Rhodococcus, and Salinispora species); plant symbionts (e.g., Frankia spp.); and insect, plant, or animal pathogens (e.g., Corynebacterium, Mycobacterium or Nocardia species) [1,11,12]. They also present in extreme environments especially the psychrophilic area, for example, Antarctica terra and desert soil [13,14,15].
2.1. Soil Actinomycetes
Actinomycetes grow as hyphae-like fungi responsible for the characteristically “earthy” smell of freshly turned healthy soil. The actinomycete population is largest in the surface layer of soils and gradually decreases with depth because of their need for oxygen [16,17]. Their estimated values range from 104 to 108 cells per gram of soil. They are sensitive to acidity/low pH (optimum pH range is 6.5–8.0) and waterlogged soil conditions. These microorganisms are primarily mesophilic, thriving in temperatures between 25 °C and 30 °C. Actinomycetes play a crucial ecological role as saprophytes, actively participating in various biological processes such as organic matter recycling, bioremediation, and promoting plant growth.
Plant growth-promoting actinomycetes use both direct (e.g., producing plant hormones) and indirect (e.g., inhibitory to plant pathogens) mechanisms to influence plant growth and protection [18,19,20,21]. They impact the process of plant biomass decomposition [22] and the microbiome around the rhizosphere [20]. They are also found in salty soil samples [23]. Ahmed et al. [24] explored the saline soil microbiome for its native structure and novel genetic elements involved in osmoadaptation. Their 16S rRNA gene sequence analysis indicated the dominance of halophilic/halotolerant phylotypes affiliated with Pseudomonadota, Actinomycetota, Gemmatimonadota, Bacteroidota, Bacillota, and Acidobacteriota. Their microbiome analysis revealed that the abundance of Actinomycetota among the other phyla was 21%.
2.2. Endophytic Actinomycetes
Endophytic actinomycetes that reside within the interior tissues of healthy plants without adversely affecting the host plant are an excellent source of potential new bioactive compounds [10,12]. Endophytic strains have been isolated from diverse plants, such as crop plants, medicinal plants, halophytes, and some woody tree species [10,25]. Examples of endophytic actinomycetes include Actinoallomurus, Actinoplanes, Allonocardiopsis, Amycolatopsis, Blastococcus, Glycomyces, Kibdelosporangium, Micrococcus, Micromonospora, Modestobacter, Nocardia, Nocardioides, Nonomuraea, Plantactinospora, Pseudonocardia, Pseudonocardia, Rothia, Saccharopolyspora, Solirubrobacter, Sphaerisporangium, Streptomyces, Streptosporangium, Wangella, and Xiangella [12].
A 16S rRNA analysis by Janso and Carter [26] categorised 113 isolated actinomycetes from 256 tissue samples (e.g., leaves, roots, and stems) collected from 113 plants. They found six families: Streptosporangiaceae (40%), Streptomycetaceae (27%), Thermomonosporaceae (16%), Micromonosporaceae (8%), Pseudonocardiaceae (8%), and Actinosynnemataceae (2%). Their findings indicate that rare actinomycetes (non-Streptomyces) predominate in plant samples. In general, the appearance of isolated, rare actinomycetes from soil samples will be either inhibited or hindered by fast-growing Streptomyces strains [27]. However, this is not the case in plant roots where the ratio of Streptomyces is low. Therefore, plant roots are an excellent potential source of rare actinomycetes and, probably, new secondary metabolites.
2.3. Actinomycetes in Compost
During the composting process, microorganisms (bacteria, actinomycetes, and fungi) are vital for organic matter degradation to produce carbon dioxide, water, heat, and humus and the relatively stable organic product as byproducts [28]. Composting generally proceeds through three main phases: (i) the mesophilic phase, (ii) the thermophilic phase, and, finally, (iii) the cooling and maturation phase. Different microbial consortia play a role during each composting phase [29]. Early decomposition is performed by mesophilic microbes, which rapidly break down soluble, readily degradable compounds. The accumulative heat causes the compost temperature to rise rapidly. As the temperature increases > 40 °C, the mesophilic microorganisms are less active and are replaced by thermophilic microbes. During the thermophilic phase, high temperatures accelerate the degradation of organic molecules such as proteins, fats, and complex carbohydrates in plant-like cellulose, hemicellulose, and lignin [30]. As these complex compounds are used up, the temperature of the compost gradually decreases, and mesophilic microorganisms once again predominate the final phase of ‘curing’ or maturation of the remaining organic matter.
Actinomycetes are typically present in the compost, particularly in the thermophilic and curing stages [31,32]. Thermophilic actinomycetes are well-known components of composts’ microflora and play an important role in habitats where organic matter decomposition occurs at elevated temperatures and under aerobic conditions (e.g., improperly stored hay, cereal grains, manure, straw, and various composts). Thermophilic actinomycetes include various genera, such as Saccharomonospora, Saccharopolyspora, Streptomyces, Thermoactinomyces, Thermobifida, and Thermomonospora.
2.4. Marine Actinomycetes
Marine habitats are a rich source of diverse and mostly uncharacterised actinomycetes. Marine habitats include coastal, deep-sea sediment, seawater, and mangrove forests. Mangrove forests are highly dynamic ecosystems that cover 75% of the world’s tropical climate, and the diversity of mangrove organisms remains less unexplored. Mangrove forests are a unique environment because they fluctuate with salinity and tidal gradients that favour their microorganisms to produce unusual metabolites [33].
Reports on marine actinomycetes have emerged since the late 19th century. Since 1980, biotechnology initially provided direction to study marine microorganisms for various applications such as drug development [34]. This study was ongoing until 1984 and used various techniques, identifying the first marine actinomycete, Rhodococcus marinonascens [11]. In 2005, the first seawater-obligate marine actinomycetes genus, Salinispora, was described. Salinispora tropica and Salinispora arenicola are novel species belonging to the family Micromonosporaceae. This genus required seawater for its growth [35]. Fencial and Jensen [36] detected novel secondary metabolites produced by Salinispora, leading to a further search for new groups of marine actinomycetes [34]. Studied marine actinomycete genera include Dietzia, Rhodococcus, Streptomyces, Salinispora, and Micromonospora. Microbiomes or traditional bacterial enumeration have been studied in marine ecosystems, including seawater [37], coral reefs [38,39,40], and mangroves [36,41,42,43]. There are various groups of microorganisms in mangrove sediments, such as Pseudomonadota, Actinomycetota, and fungi. Both Streptomyces and Micromonospora from marine habitats are good candidates for isolating potent growth-inhibiting compounds and antitumor agents. Its secondary metabolites show diverse bioactivities, such as antifungal, antitumor, and antibacterial [44]. Marine actinomycetes are one potential marine organism that can produce efficient anticancer agents such as salinosporamide A (S. tropica), actinomycin D (Streptomyces parvulus), mitomycin C (Streptomyces caespitosus), and Rakicidin D (Streptomyces sp. MWW064) [45,46].
3. Taxonomy and Classification
Early taxonomic systems for actinomycetes relied primarily on phenotypic traits, using similarities and differences in colour (spores, substrate mycelium, and soluble pigments) and melanin pigment production [47,48]. While morphological characteristics are important for describing taxa, they cannot differentiate between many genera. Polyphasic taxonomy was introduced to provide stable nomenclature and reliable identification [49]. The polyphasic approaches are based on integrating genotypic, phenotypic, and chemotaxonomic data obtained using several techniques. Currently, genotypic methods examine nucleic acids (DNA or RNA), while chemotaxonomic methods examine proteins, phenotypic features, and observable characteristics. Each method has a specific taxonomic resolution, offering different discriminatory powers at a particular level in the hierarchy. While a broad range of techniques can be readily performed, not all must necessarily be harnessed. For example, some methods for studying DNA, such as DNA base composition analysis and DNA–DNA hybridisation (DHH), are widely used to characterise bacteria. However, information from other arduous, time-consuming, technologically challenging methods, such as amino acid sequencing, is practical and only needed for a restricted number of taxa [50].
An updated actinomycete taxonomy is based on 16S rRNA- and whole genome-based phylogenetic trees. DDH values have been used for determining relatedness between closely related bacterial species. It is an available technique that offers a genuinely genome-wide distance between organisms. Wayne et al. [51] proposed a DDH value of 70% as the recommended threshold for related species, with a value of ≤70% indicating that the assayed organism belongs to a different species from the reference strains. Genomic similarity can be interpreted from the melting temperature by calculating a digital DDH (dDDH). This genome-based method was expanded by establishing several approaches in addition to BLAST to analyse high-scoring segment pairs, which are used to calculate the average nucleotide identity (ANI) and the conserved percentage of DNA. This technique correlates better with 16S rRNA gene sequences than DDH values and has a lower error ratio. The boundaries of ANI and dDDH values acceptable to propose new species are 95~96% and 70%, respectively [52].
Furthermore, molecular taxonomy, microscopic morphology and chemotaxonomy are essential characteristics to investigate the taxonomy of actinomycetes at the family and genus levels. These characteristics generally relate to the structure of the cell wall and the whole-cell sugar distribution. However, phospholipid composition and menaquinone type may also be considered for fine-tuning purposes [53]. Actinomycetes have diverse morphologies, differing mainly in the presence or absence of a substrate mycelium or aerial mycelium, the colour of the mycelium, the production of diffusible melanoid pigments, and the structure and appearance of the spores [1]. These characteristics are useful for their classification. From the above combination approaches, our group has discovered many new species of rare actinomycetes from diverse habitats, such as Actinokineospora bankokensis [54], Saccharopolyspora rhizosphaerae [55], Streptosporangium jomthongense [56], Saccharomonospora colocasiae [57] from rhizosphere soil, Amycolatopsis iheyensis [48], and Micromonospora pelagivivens [58] from marine sediments.
4. A Crucial Secondary Metabolite Producer
Actinomycetes are estimated to be the primary producer of antibiotics among all microbes and produce ~55% of all established antibiotics. Approximately 75% of these were discovered from Streptomyces, and the remaining 25% from non-Streptomyces species. Of the 22,000 known microbial secondary metabolites, 70% are produced by actinomycetes, 20% by fungi, 7% by Bacillus spp., and 1–2% by other bacteria [59].
Antibiotic production by streptomycetes is usually accumulated in a growth-phase-dependent manner. Typically, antibiotics from streptomycetes are produced in small amounts at the transition phase of conidial development when vegetative mycelium growth is slowing due to nutrient exhaustion and aerial mycelium development at the expense of nutrients released by breaking down the vegetative hyphae [60]. Typically, actinomycete secondary metabolites are structurally biosynthesised by polyketide synthase (PKS) and/or non-ribosomal peptide synthetase (NRPS). The genes governing the biosynthesis of secondary metabolites are generally clustered, with secondary metabolites synthesised from their precursors through multiple-step biosynthetic pathways [61].
The secondary metabolites of marine actinomycetes can be classified based on their chemical structure as alkaloids, peptides, polyketides, caprolactones, butanolides, polycyclic xanthones, trioxacarcins, and others. Below, we attempt to summarise the most recent findings for each bioactivity from actinomycetes from between 2016 to 2022, from the PubMed and Web of Science databases.
5. Biological Activity of Secondary Metabolites from Actinomycetes and Their Applications
5.1. Antibacterial Agents
According to the World Health Organization, the emergence of multidrug resistance by various bacterial pathogens is currently one of the biggest threats to global health and food security [62]. Therefore, finding new antibiotics remains important for expanding alternative metabolites to fight against these bacterial pathogens. Diverse actinomycete antibiotics are listed according to each class such as β-Lactams (penicillin, cephalosporines, carbapenems, monobactam, and Penicillin Binding Protein (PBP) inhibitors, ansamycines, macrolides, tetracyclines and aminoglycosides and antibiotic peptides [63].
Since 2010, our research group has discovered actinomycin production by the rhizosphere Streptomyces sp. SJE177 with activity against multidrug-resistant Staphylococcus aureus [64] and successfully purified decatromicin derivatives with anti-Clostidium activity from rhizosphere Actinomadura sp. 2EPS [65]. Currently, more antibacterial metabolites were further discovered from actinomycetes. Table 1 shows that most antibacterial metabolites were identified from Streptomyces. The remaining were found from rare actinomycetes from the genera Actinoallourus, Actinomadura, Amycolatopsis, Kibdelosporangium, Kitasatospora, Kocuria, Microbacterium, Micromonospora, Rhodococcus, Nocardiopsis, Pseudonocardia, Streptomonospora, Streptosporangium, Thermoactinomyces, and Verrucosispora.
5.2. Antifungal Agents
Among the different types of drugs on the market, antifungal antibiotics, which have an important role in controlling fungal infections, are relatively small but significant groups of drugs. Antifungal agents are widely applied in humans, medicine, agriculture, and veterinary medicine. There are five major classes of antifungal compounds: (i) polyene antibiotics, (ii) allylamines and thiocarbamates, (iii) azole derivatives, (iv) morpholines, and (v) nucleoside analogues [154]. In the late 1950s, the first polyene macrolide antifungal antibiotic from Streptomyces species was discovered [155,156]. Polyene antifungal compounds such as amphotericin B are the standard therapy for fungal infections. The mode of action of amphotericin B is its interaction with membrane sterol, which results in aqueous pore production with the polyene hydroxyl residues facing inward, leading to altered permeability, leakage of vital cytoplasmic components, and the death of the organism [157]. The azole, allylamine, and thiocarbamate classes target ergosterol, the same as polyene antibiotics. The morpholine class inhibits sterol synthesis, and the nucleoside analogue class targets DNA synthesis [154]. Most recent antifungal metabolites were identified from Streptomyces. The remaining were found in rare actinomycetes from the genera Actinomadura, Amycolatopsis, Actinokineospora, Norcardia, Pseudonocardia, Saccharothrix, and Umezawaea (Table 2). Our research group published the antifungal activities of bafilomycin from the rhizosphere Streptomyces sp. SBI034 [158] and thailandins from Actinokineospora bangkokensis strain 44EHWT [159] against Colletotrichum spp., the causative agent of anthracnose disease. Another study showed that adding a halogenated benzoyl group to the benzoate derivative’s 3-OH group improved altholactone’s antifungal activity against Cryptococcus neoformans and Saccharomyces cerevisiae [160].
5.3. Immunosuppressive Agent
Immunosuppressants are essential drugs that significantly decrease the risks of rejecting a transplanted organ. In addition, immunosuppressant drugs are also used to treat many autoimmune disorders, such as Crohn’s disease (chronic inflammation of the digestive tract), rheumatoid arthritis, and patchy hair loss (alopecia areata). Since 1970, the peptide antibiotic cyclosporine A has been the primary immunosuppressant used in transplantation. Various immunosuppressant drugs originally isolated as antifungal antibiotics are produced by microorganisms, such as ascomycin from Streptomyces hygroscopicus and tacrolimus from Streptomyces tsukubaensis [186]. Regarding their mode of action, immunosuppressant drugs act by binding to immunophilin, which is involved in T-cell activation and proliferation [187].
5.4. Biocontrol Agents
Global attempts to discover natural products as biocontrol agents for plant protection have notably increased. Being the most proactive, Streptomyces appear to be a readily available natural choice for finding new ways to combat plant pathogens and show appreciable biocontrol action against diverse phytopathogens. However, only a few have been developed as commercial products for plant applications in agriculture. Table 3 lists the microbial pesticides registered in countries worldwide.
Actinomycetes are known for improving compost quality and increasing its nutrient content. They also increase the odour of compost since they can completely digest the organic matter present in compost [188]. It has been shown that the thermophilic Actinomycetota Streptomyces sp. No. 101 and Micromonospora sp. No. 604 can completely degrade yeast debris and sanitise the compost [189,190]. In addition, Streptomyces thermodiastaticus was found to produce various extracellular enzymes involved in pathogenic yeast cell lysis, such as Candida albicans. Some thermophilic Actinomycetota can suppress plant diseases and thereby promote good crop plant health, increasing crop yields. Therefore, these thermotolerant Actinomycetota could be used as an alternative to commercial pesticides.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 3.
List of Streptomyces spp.—based products as biocontrol agents available in the market worldwide [191].
Table 3.
List of Streptomyces spp.—based products as biocontrol agents available in the market worldwide [191].
| Commercial Product Name | Organism As Active Substance | Registered As a Microbial Pesticide | Targeted Pest/Pathogen/Disease |
|---|---|---|---|
| Actinovate, Novozymes BioAg Inc., Milwaukee, WI, USA | S. lydicus WYEC 108 | Canada, USA | Soilborne diseases, viz. Pythium, Fusarium, Phytophthora, Rhizoctonia, and Verticillium; foliar diseases such as powdery and downy mildew, Botrytis, Alternaria, Postia, Geotrichum, and Sclerotinia |
| Mycostop, Verdera Oy, Espoo, Finland | Streptomyces K61 | EU, Canada, USA | Damping off caused by Alternaria, R. solani, Fusarium, Phytophthora, Pythium wilt, and root diseases |
| Mykocide, KIBC Co., Ltd., Yongin, Gyeonggi-do, Republic of Korea | S. colombiensis | Republic of Korea | Powdery mildews, grey mold, and brown patch |
| Bactophil | Streptomyces albus | Ukraine | Seed germination diseases |
5.5. Antitumor Compounds
The novel antitumor drug discovery tends to be accomplished by natural product-producing marine microorganisms. Marine-derived actinomycetes are one of the promising candidates in antitumor compound screening. From 2007 to 2017, Micromonospora, Salinispora, and Verrucosispora were among the top new secondary metabolite producers [192]. In previous publications, the reported compounds were categorised into four groups based on natural product classes: polyketides, alkaloids, peptides, and quinones. Actinomycetes have notable anticancer therapeutic potential, especially those whose products are associated with minimal side effects compared to conventional chemotherapy, such as salinosporamide A [193]. Adriamycin, isolated from Streptomyces peucetius [194], inhibits DNA replication and is an anticancer drug. Other effective products for cancer chemotherapeutics are actinomycin D, bleomycin, anthracyclines (daunorubicin), and mitosanes (mitomycin C). These drugs were obtained from Streptomyces verticillus, Streptomyces peucetius, S. caespitosus, and other intrageneric isolates [195]. Marine Actinomycetota compounds with antitumor potential include streptochlorin, actinofuranones, aureoverticillactam, chalocomycin B, cyanosporasides, komodoquinones, nonactin, resitoflavine, sporolides, tetracenomycin D, thiocoraline, t-muurolol, butenolides, echinosporins, rakicidin D, and streptokordin [45,195,196]. Important secondary metabolites from marine actinomycetes with antitumor potential include streptopyrrolidine, cyclo-(l-Pro-l-Met), streptochlorin, lynamicins, marizomib, and thiocoraline [195]. Two examples of novel anticancer metabolites are the compound extracts ULDF4 and ULDF5 derived from Streptomyces strains found in Lagos, Nigeria. ULDF4 and ULDF5 show cytotoxicity against human acute myelocytic leukaemia, cervical carcinoma, human gastric carcinoma, breast adenocarcinoma, and human acute promyelocytic leukaemia. ULDF4 and ULDF5 are structurally similar to staurosporine and kigamicin, compounds known to induce apoptosis and necrosis, respectively. Ketomycin is another prospective antitumor compound. Ketomycin suppressed breast carcinoma cell migration and invasion, inhibited nuclear factor kappa-B (NF-κB) activity in upstream signalling by impeding the autophosphorylation of inhibitory-κB kinases alpha (IKK-α) and beta (IKK-β), and minimised the 3D invasion of breast carcinoma cells at nontoxic concentrations [197]. Therefore, ketomycin is an effective antibiotic and a structurally simple antitumor agent for mammalian cells. The search for further antitumor agents also includes the analysis of biosynthetic (BGCs) and chemotherapeutic (CGCs) gene clusters. Complementary anticancer treatments have been discovered in Streptomyces via the diverse and variable patterns of the phylogenetic distribution of BGCs and CGCs [195]. These hybrid BGCs and CGCs are prospective sources of novel secondary metabolites and chemotherapeutic agents for pharmaceuticals. The use of Streptomyces compounds such as staurosporine, kigamicin, and ketomycin and BGCs/CGCs should be further investigated for developing new antitumor treatments.
Most recent anticancer metabolites were identified from Streptomyces. The remaining were found in rare actinomycetes from the genera Actinoalloteichus, Actinokineospora, Actinomadura, Actinosynnema, Amycolatopsis, Catenuloplanes, Dietzia, Microbacterium, Micromonospora, Nocardiopsis, Nocardiopsis, Nonomuraea, Saccharomonospora, Tsukamurella, Umezawaea, and Verrucosispora (Table 4).
5.6. Antiviral Agents
The outbreaks of the influenza A virus (IAV), severe acute respiratory syndrome coronavirus (SARS-CoV), Middle East respiratory syndrome coronavirus, and, recently, SARS-CoV-2 highlight the need for discovering effective antiviral drugs against respiratory RNA virus. Indeed, the coronavirus disease 2019 caused a global pandemic with high mortality rates worldwide. Natural products derived from microbial sources are still an excellent structural motif for discovering new therapeutics, including antiviral agents [268]. About 50% of the US Food and Drug Administration (FDA)-approved natural product-based drugs are of microbial origin, notably antivirals [269]. Vidarabine (Ara-A) isolated from Streptomyces antibioticus was one of the earliest antiviral nucleoside analogues [270]. In addition, several ansamycin antibiotics have shown antiviral properties against diverse infectious [271]. The study of Raveh et al. purified a novel metabolite (an Antimycin A derivative) with significant antiviral activity from Streptomyces kaviengensis [272]. This compound showed activity against the Western equine encephalitis virus with a half-maximal inhibitory concentration (IC50) of <4 nM. The mode of action revealed the disruption of mitochondrial electron transport and pyrimidine biosynthesis [273]. Streptomyces citricolor produced aristeromycin that showed potent anti-IAV activity [274]. Protease inhibitor PISC-2002 from the culture supernatant of Streptomyces chromofuscus showed antiviral activity against influenza virus A/Rostock/34 (H7N7) [275]. The microbial-derived FDA-approved anti-parasitic drug ivermectin, a semisynthetic pentacyclic sixteen-membered lactone derived from the soil bacterium Streptomyces avermitilis, was recently reported. Ivermectin was shown to be an effective in vitro inhibitor of SARS-CoV-2 replication [276,277].
Most recent antiviral metabolites were identified from Streptomyces, with the others identified from Kutzneria (Table 5). Our research group discovered the in vitro antiviral activity of spectrotetronates and decatromicins against Dengue virus serotype 2, with a suggested mode of action in preventing viral replication and assembly via inhibiting the viral protease [278].
5.7. Other Activities
Our research group also discovered bioactive compounds with other activities: adipocyte differentiation, anti-trypanosomal activity, and androgen receptor (AR) antagonistic activity. Three modified amino acids (jomthonic acids A-C) were isolated from a soil-derived Streptomyces sp. BB47, with jomthonic acids A and B inducing adipocyte differentiation [289,290]. Five new anti-trypanosomal macrolides (actinoallolides) were identified from the cultured broth of Actinoallomurus fulvus MK10-036. Actinoallolide A showed the best in vitro anti-trypanosomal activity with an IC50 of 0.0049 μg mL–1 without cytotoxicity [291]. Antarlides A-E were isolated from Streptomyces sp. BB47. Antarlide B inhibited the transcriptional activity of wildtype and mutant ARs and could be used as an AR antagonist. In 2020, norditerpenoid k4610422, an inhibitor of testosterone-5α reductase, was identified from Actinomadura spp. [292].
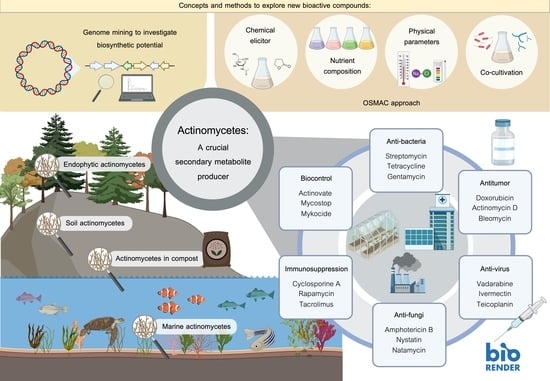
6. Concepts and Methods to Explore New Bioactive Compounds
Many new antibiotics were identified in massive screening programs during the ‘golden age’ of antibiotic discovery from the late 1940s to the late 1960s. While the supply of new antibiotics has declined over the last decade, the emergence of multidrug-resistant microbial pathogens has increased [293,294]. Therefore, finding new therapeutic active compounds remains of scientific interest.
6.1. Exploring New Habitats or Extreme Environments As A Source for Novel Strains
Over the past 50 years, researchers have investigated antibiotics and metabolites from terrestrial microorganisms, but 99% of their metabolites are known compounds [192]. Therefore, the chance of discovering novel secondary metabolites has diminished. Currently, only ~1% of existing actinomycetes can be identified based on DNA analyses. However, actual microbial diversity is much greater than expected [295]. The focus of industrial screening has moved to markers of less exploited genera of rare actinomycetes (e.g., Actinokineospora, Jishengella, Micromonospora, Nocardiopsis, Salinispora, Saccharomonospora, Saccharopolyspora, and Verrucosispora) [296]. Therefore, the isolation of novel species from diverse ecosystems may result in new unprecedented bioactive compounds produced by them.
Extremophile organisms live in extreme habitats and have unique survival mechanisms to withstand harsh conditions, such as high temperature, extreme pH, salinity, pressure, and aridity [297]. Exploration of unique ecological niches and new isolation methods for novel genera/species of actinomycetes may identify new BGCs. While actinomycetes are commonly present in terrestrial and aquatic ecosystems, studies have focused on isolating them from extreme geographical locations, including oceans [40,93,109,206,298], hot springs [299,300], desert soil [13,14,116,301,302], salt lakes [23,303]. Among these extreme habitats, deserts and the deep sea were the most favourable environments for isolating bioactive compounds with potential applications in medicine from actinomycetes [304]. Marine habitats have received increasing attention in recent decades because the ocean covers > 70% of the earth’s surface and has a unique ecosystem. Marine habitats show extreme light, oxygenation, and temperature conditions. Therefore, microorganisms could adapt to survive in these extreme environments and produce unique secondary metabolites [46]. One of the most important sources of new bioactive compounds is endophytic actinomycetes. The association of actinomycetes with plants constitutes a unique trait conferring specific biological and chemical features. They can potentially produce various valuable metabolites [10,12,18,25,135,191,305].
6.2. Genome Mining to Investigate Biosynthetic Potential
Genomic-guided approaches help discover new natural products leading to an increasing number of complete genome sequence projects. The publication of the first complete genome sequence of Streptomyces coelicolor showed surprisingly more suspected biosynthetic clusters than expected, indicating that many new bioactive compounds remain to be discovered and exploited [306,307]. Next-generation sequencing has revolutionised the field and dramatically increased the reported genomes for actinomycetes. The complete annotated genome sequences of antibiotic-producing actinomycetes, including S. avermitilis, Saccharopolyspora erythraea, S. tropica, and Rhodococcus sp. RHA1 revealed numerous (≥20 potential secondary metabolites) PKS, NRPS, and many other small-molecule biosynthetic pathways [295]. This information indicates that actinomycetes remain promising sources of novel bioactive compounds.
Genome mining primarily aims to identify biosynthetic pathways that produce novel bioactive molecules. Clusters of silent or cryptic biosynthetic genes, usually associated with a known chemical class such as polyketide and NRPS, are scanned in silico to search for consensus motifs and predict possible chemical structures. Developing rapid automatic search algorithms is essential to take full advantage of genome sequence information. Currently, the most widely used software to identify such gene clusters is Antibiotics and Secondary Metabolite Analysis Shell (antiSMASH; https://antismash.secondarymetabolites.org, accessed on 27 July 2023) [308]. Other computational tools, such as CLUSEAN [309], PRISM [310], NP.searcher [311], and ClustScan [312], have been developed to identify secondary metabolite BGCs (smBGCs) within the genome. These bioinformatics tools rely on the highly conserved sequences within the smBGCs to map their location. However, the predicted smBGCs still require extensive laboratory work, such as activating silenced smBGCs and purifying and elucidating potential compounds [313].
The mining of whole genomes from actinomycete species reveals a great genetic potential to synthesise secondary metabolites, including antibiotics. However, only some of these clusters are expressed under standard laboratory conditions. Several methods have been developed to trigger the expression of ‘silent gene clusters’, including optimising fermentation conditions, inducing mutations in ribosomal proteins or RNA polymerase, co-cultivating the antibiotic producer with other microorganisms, and activating or disrupting pathway-specific controlling gene clusters (Figure 3). If a gene cluster of interest cannot be expressed in its original host, it must be cloned into a suitable vector and expressed in a heterologous host. A heterologous expression established an approach for unlocking silent or cryptic gene clusters [314]. Several techniques for assembling and cloning of BGCs have been developed and optimised. Generally, this method includes three steps: (i) cloning of the target BGCs, (ii) engineering of the target BGCs, and (iii) transformation into the selected heterologous host. For cloning large BGCs, a genomic library is constructed by cosmid or fosmid. Transformation-associated recombination is an alternative used for direct cloning of BGCs, where large BGCs can be assembled from multiple overlapping PCR products or restriction fragments isolated from genomic libraries. A silent biosynthetic pathway generating a non-ribosomal peptide from Saccharomonospora sp. CNQ-490 was successfully cloned and functionally expressed in S. coelicolor. For example, expressing the terpene synthase gene from S. avermitilis in an Escherichia coli host resulted in the synthesis of a novel tricyclic sesquiterpene: avermitilol [315]. Our research group is working on genome mining of Actinokineospora bangkokensis 44EHWT to further investigate bioactive metabolites in addition to thailandins [159].
6.3. OSMAC Approach
In One Strain-Many Compounds (OSMAC), new approaches unlocking the silent gene clusters are used to improve secondary metabolite production in microorganisms through changes in growth conditions. The name was coined by Zeeck et al. [316]. Its basis is that a single strain of microbes can biosynthesise different compounds when growing under particular conditions. Romano et al. [317] reported that OSMAC strategies are changes in nutrient regimes, physical parameters (e.g., temperature), co-cultivation, and other environmental cues (Figure 4). Variation of the medium is a simple but successful strategy since it directly affects microbial metabolism and differentiation. Changes in nutrient composition, C/N ratio, metal ions, and salt content influence gene expression in the microbial culture. Additionally, a lack of nutrients may induce microbes to attempt to survive by inducing special functional genes. Co-cultivation with other strains may create competitive or favourable conditions. The co-cultivation method possibly alters secondary metabolite production of both known and novel compounds from the silent gene clusters. Mixed cultivation of two strains of fungi (Nos. 1924 and 3893) produced one novel 1-isoquinolone analogue and its methyl ester that was not found with isolated cultivation. Co-cultivation with mycolic acid-containing bacteria was also shown to produce new secondary metabolites in Streptomyces [318]. Using γ-butyrolactone (GBL) as a chemical elicitor can induce the biosynthesis of antibiotics in actinomycetes [319]. N-acetylglucosamine (GlcNAc) also effectively stimulated bioactive compound production in Actinokineospora spheciospongiae sponge-derived actinomycetes [320]. Additionally, the co-cultivation of actinomycetes with fungi led to new compound production: borrelidins J and K and 7-methoxy-2,3-dimethylchromone-4-one [321].
6.4. Co-Cultivation Technique
Co-cultivation is the cultivation of two or more microbe strains. In nature, microbes live together in a community. When cultured with other microbes, they may produce a new compound because of nutrient competition or friendly relationships encouraging them to. Additionally, co-cultivation can increase the yield of existing metabolites. For example, the co-culturing of marine Streptomyces with human pathogens can induce the production of tree antibiotics and improve activity against Gram-positive human pathogens. Co-culturing of Streptomyces rochei MB307, actinomycetes, and the fungi Rhinocladiella similis 35 produced new metabolites [321]. A red pigment was produced by Streptomyces lividans TK23 during co-cultivation with Tsukamurella pulmonis TP-B0596. Moreover, the co-culturing of rare actinomycetes Umezawaea sp. RD066910 with mycolic acid-containing bacteria Tsukamurella pulmonis TP-B0596 induced the production of umezawamides A and B, new macrolactams toxic to P388 murine leukaemia cells, while umezawamides A can also inhibit Candida albicans [185].
6.5. Using Chemical Elicitors
Actinomycetes employ renowned extracellular signal molecules such as N-acetylglucosamine and γ-butyrolactone to trigger the activation of cryptic secondary metabolism gene operons. The utilisation of chemical elicitors represents a potent approach capable of unlocking the expression of cryptic biosynthetic gene clusters in actinomycetes. Through this method, the production of new or low-yield secondary metabolites is achieved, thereby broadening the diversity, and enhancing the potential for discovering novel bioactive compounds with diverse biological activities [320,322,323]. The review delves into various types of chemical elicitors and their effects on secondary metabolites.
6.5.1. γ-. Butyrolactones and Related Regulators
Organic and low-molecular-weight gamma butyrolactones (GBLs) are chemical agents widely used to trigger secondary metabolite production because bacteria in the Streptomyces genus produce them as microbial hormones (autoregulators). These hormones control secondary metabolite production and bacterial morphological changes. GBLs’ major backbone is 2-(1″-oxo or 1 ″hydroxyalkyl)-3-hydroxymethyl butyrolactone. GBLs are classified into three major types based on the characteristic of their side chain: the A-factor type (1″-keto-type), the virginiae butanolide type ((1 ″S)-hydroxy-type), and the IM-2 type ((1 ″R)-hydroxy-type). Choi et al. described that Streptomyces and rare actinomycetes possess GBL autoregulators, including Actinoplanes teichomyceticus and Amycolatopsis mediterranei [324]. Examples of GBL regulation include the production of virginiamycin by Streptomyces virginiae, an antibiotic used in the ethanol industry to eliminate bacteria; the production of showdomycin and minimycin by Streptomyces lavendulae; and the production of pigmented antibiotics Act and Red by S. coelicolor. In previous studies, it was demonstrated that the addition of an exogenous butyrolactone compound led to a significant increase in validamicyn antibiotic production in Streptomyces hygroscopicus 5008 [322].
However, in actinomycetes, GBLs are not the sole regulators present. Other types of autoregulators have been reported, such as the pi-factor from S. natalensis and l-N-methylphenylalanyl-dehydrobutyrine diketopiperazine from S. globisporus [322]. Goadsporin, a potent 19-amino acid peptide discovered in the culture broth of Streptomyces sp. TP-A0584 exhibits remarkably broad elicitation activity on actinomycetota. Goadsporin has the capability to stimulate S. lividans to produce red pigment and facilitate the process of sporulation [325].
6.5.2. N-Acetylglucosamine (GlcNAc)
GlcNAc is a monosaccharide glucose amide derivative, the secondary amide of glucosamine and acetic acid. It is a component of a biopolymer in the bacterial cell wall, peptidoglycan layer, or murein. GlcNAc is also a chitin monomer, a polymer in the exoskeletons of arthropods, radula in molluscs, and fungi cell walls. Tawfike et al. reported that GlcNAc could activate cryptic gene clusters in sponge-derived rare actinomycete Actinokineospora spheciospongiae sp. during cultivation in broth and on solid agar. Four new actinosporins (E–H) were discovered in broth culture. Additionally, fridamycins H and I and actinosporins C, D, and G were produced in solid agar culture [320]. Moreover, their previous study explored the effect of GlcNAc on cultured Micromonospora sp. RV43, Rhodococcus sp. RV157, and Actinokineospora sp. EG49. They found that GlcNAc-induced Micromonospora sp. RV43 to produce 3-formylindole and guaymasol, Rhodococcus sp. RV157 to produce the siderophore bacillibactin antibiotic surfactin and Actinokineospora sp. EG49 to increasingly produce actinosporins E–H [326].
7. Future Perspectives on Actinomycetes
Actinomycetes produce diverse bioactive metabolites, many of which play important roles as therapeutic lead compounds. Therefore, their exploration can provide an enormous reservoir of potentially active compounds [63,327,328]. Actinomycetes are distributed across diverse habitats, including terrestrial and marine ecosystems. They can be isolated from soil, water, sediments, plants, and insects [38,329,330,331]. While they already provide various therapeutic drugs available on the commercial market, many members of this bacterial group and, importantly, their potential secondary metabolites with possible immense therapeutic values remain underexplored [328]. New genera and species are generally promising sources of novel bioactive secondary metabolites. Therefore, it is crucial to understand the strains’ biodiversity, to further investigate and identify new genera or species, and to optimise compound production processes [3,4,296]. An investigative approach to screen and select new strains with a potential therapeutic metabolite should be considered and used. In addition, many BGCs are usually found in their genome sequence, and many clusters are silenced. Activating these silent gene clusters may lead to new compound discovery [332,333]. The preliminary identification of the whole genome sequence can help to investigate the biodiversity of strains in various ecosystems and identify their BGCs. Genome-dependent mining can be used to identify the silent BGCs in their analysed genome sequences [334]. Genetic engineering by cloning and expressing the whole BGC in a suitable heterologous host could be an alternative method to activate these silent gene clusters [314,335,336,337]. It is known that one bacterium can produce new metabolites in the presence of other bacteria or fungi. Therefore, co-cultivation approaches have been adopted, and many silent gene clusters have been successfully activated [321,338]. The competition for nutrients and any signal molecules from other microorganisms may elicit new compound production from the target host strain. However, while co-cultivation can activate silent gene clusters, it can sometimes silence others. Therefore, the selection of co-cultivation partners is challenging. A better understanding and elucidation of the key signal in eliciting silent gene cluster expression is necessary to obtain new potential metabolites.
8. Conclusions
Despite the huge amount of available research information on exploring actinomycetes and their secondary metabolites, there remains significant potential for discovering new bioactive agents from these microorganisms. Integrative approaches involving new cultivation techniques, chemical elicitor induction, genome mining, and gene cluster activation should be employed to find new compounds with essential functions. Rapid progress in using genetic tools and several approaches for isolating new compounds with novel bioactivity will encourage researchers in this field to obtain promising candidates for fighting against emerging pathogens and various diseases, including non-communicable diseases. Novel genera and novel species of actinomycetes from diverse habitats are still waiting to be discovered. Marine habitats represent an underexplored ecosystem rich in microbial and compound diversity, making their metabolites highly promising for commercial applications. While marine actinomycetes may produce some of the same metabolites as their terrestrial counterparts, they have also demonstrated to be a valuable source of potential novel metabolites, especially in the case of rare marine actinomycetes.
Author Contributions
Conceptualization, B.I., J.E. and W.P.; writing—original draft preparation, B.I., C.N., N.C., J.E. and W.P.; writing—review and editing, J.E., B.I. and W.P.; visualization, C.N., N.C. and B.I. supervision, W.P.; funding acquisition, B.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding. The APC was funded by Mahidol University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors would like to thank T. Pemberton for proofreading the manuscript and Thanikarn Udomchalothorn for facilitating the visualization.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.P.; Clement, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; The John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Tiwari, K.; Gupta, R.K. Rare actinomycetes: A potential storehouse for novel antibiotics. Crit. Rev. Biotechnol. 2012, 32, 108–132. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Yang, L.-J.; Zhang, W.-D.; Shen, Y.-H. The secondary metabolites of rare actinomycetes: Chemistry and bioactivity. RSC Adv. 2019, 9, 21964–21988. [Google Scholar] [CrossRef] [PubMed]
- Al-Fadhli, A.A.; Threadgill, M.D.; Mohammed, F.; Sibley, P.; Al-Ariqi, W.; Parveen, I. Macrolides from rare actinomycetes: Structures and bioactivities. Int. J. Antimicrob. Agents 2022, 59, 106523. [Google Scholar] [CrossRef] [PubMed]
- Prudence, S.M.M.; Addington, E.; Castaño-Espriu, L.; Mark, D.R.; Pintor-Escobar, L.; Russell, A.H.; McLean, T.C. Advances in actinomycete research: An ActinoBase review of 2019. Microbiology 2020, 166, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Qinyuan, L.; Xiu, C.; Yi, J.; Chenglin, J. Morphological Identification of Actinobacteria. In Actinobacteria; Dharumadurai, D., Yi, J., Eds.; IntechOpen: Rijeka, Croatia, 2016; pp. 59–86. [Google Scholar]
- Goodfellow, M.; Williams, S.T. Ecology of actinomycetes. Annu. Rev. Microbiol. 1983, 37, 189–216. [Google Scholar] [CrossRef]
- van der Meij, A.; Worsley, S.F.; Hutchings, M.I.; van Wezel, G.P. Chemical ecology of antibiotic production by actinomycetes. FEMS Microbiol. Rev. 2017, 41, 392–416. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, A.; Takahashi, Y. Endophytic actinomycetes: Promising source of novel bioactive compounds. J. Ant. 2017, 70, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmke, E.; Weyland, H. Rhodococcus marinonascens sp. nov., an actinomycete from the sea. Int. J. Syst. Bacteriol. 1984, 34, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Masand, M.; Jose, P.A.; Menghani, E.; Jebakumar, S.R.D. Continuing hunt for endophytic actinomycetes as a source of novel biologically active metabolites. World J. Microbiol. 2015, 31, 1863–1875. [Google Scholar] [CrossRef]
- Kumar, S.; Solanki, D.S.; Parihar, K.; Tak, A.; Gehlot, P.; Pathak, R.; Singh, S.K. Actinomycetes isolates of arid zone of Indian Thar Desert and efficacy of their bioactive compounds against human pathogenic bacteria. Biol. Futur. 2021, 72, 431–440. [Google Scholar] [CrossRef]
- Mohammadipanah, F.; Wink, J. Actinobacteria from arid and desert habitats: Diversity and biological activity. Front. Microbiol. 2016, 6, 1541. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.J.; Crevelin, E.J.; Souza, D.T.; Lacerda-Júnior, G.V.; de Oliveira, V.M.; Ruiz, A.L.T.G.; Rosa, L.H.; Moraes, L.A.B.; Melo, I.S. Actinobacteria from Antarctica as a source for anticancer discovery. Sci. Rep. 2020, 10, 13870. [Google Scholar] [CrossRef]
- Zenova, G.M.; Manucharova, N.A.; Zvyagintsev, D.G. Extremophilic and extremotolerant actinomycetes in different soil types. Eurasian Soil Sci. 2011, 44, 417–436. [Google Scholar] [CrossRef]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes benefaction role in soil and plant health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef]
- Aamir, M.; Rai, K.K.; Zehra, A.; Dubey, M.K.; Samal, S.; Yadav, M.; Upadhyay, R.S. Endophytic actinomycetes in bioactive compounds production and plant defense system. In Microbial Endophytes; Kumar, A., Singh, V.K., Eds.; Woodhead Publishing: Sawston, UK, 2020; pp. 189–229. [Google Scholar] [CrossRef]
- Chaurasia, A.; Meena, B.R.; Tripathi, A.N.; Pandey, K.K.; Rai, A.B.; Singh, B. Actinomycetes: An unexplored microorganisms for plant growth promotion and biocontrol in vegetable crops. World J. Microbiol. 2018, 34, 132. [Google Scholar] [CrossRef] [PubMed]
- Nonthakaew, N.; Panbangred, W.; Songnuan, W.; Intra, B. Plant growth-promoting properties of Streptomyces spp. isolates and their impact on mung bean plantlets’ rhizosphere microbiome. Front. Microbiol 2022, 13, 967415. [Google Scholar] [CrossRef] [PubMed]
- Intra, B.; Mungsuntisuk, I.; Nihira, T.; Igarashi, Y.; Panbangred, W. Identification of actinomycetes from plant rhizospheric soils with inhibitory activity against Colletotrichum spp., the causative agent of anthracnose disease. BMC Res. Notes 2011, 4, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, Y.; Dolfing, J.; Guo, Z.; Chen, R.; Wu, M.; Li, Z.; Lin, X.; Feng, Y. Important ecophysiological roles of non-dominant Actinobacteria in plant residue decomposition, especially in less fertile soils. Microbiome 2021, 9, 84. [Google Scholar] [CrossRef]
- Guan, T.W.; Lin, Y.J.; Ou, M.Y.; Chen, K.B. Isolation and diversity of sediment bacteria in the hypersaline aiding lake, China. PLoS ONE 2020, 15, e0236006. [Google Scholar] [CrossRef]
- Ahmed, V.; Verma, M.K.; Gupta, S.; Mandhan, V.; Chauhan, N.S. Metagenomic Profiling of Soil Microbes to Mine Salt Stress Tolerance Genes. Front. Microb. 2018, 9, 159. [Google Scholar] [CrossRef] [Green Version]
- Rungin, S.; Indananda, C.; Suttiviriya, P.; Kruasuwan, W.; Jaemsaeng, R.; Thamchaipenet, A. Plant growth enhancing effects by a siderophore-producing endophytic streptomycete isolated from a Thai jasmine rice plant (Oryza sativa L. cv. KDML105). Antonie Van Leeuwenhoek 2012, 102, 463–472. [Google Scholar] [CrossRef]
- Janso, J.E.; Carter, G.T. Biosynthetic Potential of Phylogenetically Unique Endophytic Actinomycetes from Tropical Plants. Appl. Environ. Microbiol. 2010, 76, 4377–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayakawa, M. Studies on the isolation and distribution of rare actinomycetes in soil. Actinomycetologica 2008, 22, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Hemati, A.; Nazari, M.; Asgari Lajayer, B.; Smith, D.L.; Astatkie, T. Lignocellulosics in plant cell wall and their potential biological degradation. Folia Microbiol. 2022, 67, 671–681. [Google Scholar] [CrossRef]
- Hemati, A.; Aliasgharzad, N.; Khakvar, R.; Delangiz, N.; Asgari Lajayer, B.; van Hullebusch, E.D. Bioaugmentation of thermophilic lignocellulose degrading bacteria accelerate the composting process of lignocellulosic materials. Biomass Conv. Bioref. 2022, 12, 1–15. [Google Scholar] [CrossRef]
- Hemati, A.; Aliasgharzad, N.; Khakvar, R.; Khoshmanzar, E.; Asgari Lajayer, B.; van Hullebusch, E.D. Role of lignin and thermophilic lignocellulolytic bacteria in the evolution of humification indices and enzymatic activities during compost production. Waste Manag. 2021, 119, 122–134. [Google Scholar] [CrossRef] [PubMed]
- de Gannes, V.; Eudoxie, G.; Hickey, W.J. Prokaryotic successions and diversity in composts as revealed by 454-pyrosequencing. Bioresour. Technol. 2013, 133, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Takaku, H.; Kodaira, S.; Kimoto, A.; Nashimoto, M.; Takagi, M. Microbial communities in the garbage composting with rice hull as an amendment revealed by culture-dependent and -independent approaches. J. Biosci. Bioeng. 2006, 101, 42–50. [Google Scholar] [CrossRef]
- Setyati, W.A.; Pringgenies, D.; Soenardjo, N.; Pramesti, R. Actinomycetes of secondary metabolite producers from mangrove sediments, Central Java, Indonesia. Vet. World 2021, 14, 2620. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.E.F.; Moustafa, M.S.; El-Wahed, A.A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.R.; Dwight, R.; Fenical, W. Distribution of actinomycetes in near-shore tropical marine sediments. Appl. Environ. Microbiol. 1991, 57, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.R.; Mafnas, C. Biogeography of the marine actinomycete Salinispora. Environ. Microbiol. 2006, 8, 1881–1888. [Google Scholar] [CrossRef]
- Bienhold, C.; Zinger, L.; Boetius, A.; Ramette, A. Diversity and Biogeography of Bathyal and Abyssal Seafloor Bacteria. PLoS ONE 2016, 11, e0148016. [Google Scholar] [CrossRef] [PubMed]
- Betancur, L.A.; Naranjo-Gaybor, S.J.; Vinchira-Villarraga, D.M.; Moreno-Sarmiento, N.C.; Maldonado, L.A.; Suarez-Moreno, Z.R.; Acosta-González, A.; Padilla-Gonzalez, G.F.; Puyana, M.; Castellanos, L.; et al. Marine Actinobacteria as a source of compounds for phytopathogen control: An integrative metabolic-profiling/bioactivity and taxonomical approach. PLoS ONE 2017, 12, e0170148. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, H.M.; Kalendar, A.A. Coral-associated Actinobacteria: Diversity, abundance, and biotechnological potentials. Front. Microb. 2016, 7, 204. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento-Vizcaíno, A.; González, V.; Braña, A.F.; Palacios, J.J.; Otero, L.; Fernández, J.; Molina, A.; Kulik, A.; Vázquez, F.; Acuña, J.L.; et al. Pharmacological Potential of Phylogenetically Diverse Actinobacteria Isolated from Deep-Sea Coral Ecosystems of the Submarine Avilés Canyon in the Cantabrian Sea. Microb. Ecol. 2017, 73, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ye, Y.; Wang, R.; Zhang, Y.; Wu, C.; Debnath, S.C.; Ma, Z.; Wang, J.; Wu, M. Streptomyces nigra sp. nov. Is a Novel Actinobacterium Isolated From Mangrove Soil and Exerts a Potent Antitumor Activity in Vitro. Front. Microbiol. 2018, 9, 1587. [Google Scholar] [CrossRef] [Green Version]
- Kemung, H.M.; Tan, L.T.H.; Chan, K.G.; Ser, H.L.; Law, J.W.F.; Lee, L.H.; Goh, B.H. Streptomyces sp. Strain MUSC 125 from Mangrove Soil in Malaysia with Anti-MRSA, Anti-Biofilm and Antioxidant Activities. Molecules 2020, 25, 3545. [Google Scholar] [CrossRef]
- Lin, X.; Hetharua, B.; Lin, L.; Xu, H.; Zheng, T.; He, Z.; Tian, Y. Mangrove Sediment Microbiome: Adaptive Microbial Assemblages and Their Routed Biogeochemical Processes in Yunxiao Mangrove National Nature Reserve, China. Microb. Ecol. 2019, 78, 57–69. [Google Scholar] [CrossRef]
- Xu, D.; Han, L.; Li, C.; Cao, Q.; Zhu, D.; Barrett, N.H.; Harmody, D.; Chen, J.; Zhu, H.; McCarthy, P.J.; et al. Bioprospecting deep-sea actinobacteria for novel anti-infective natural products. Front. Microbiol. 2018, 9, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olano, C.; Méndez, C.; Salas, J.A. Antitumor compounds from marine actinomycetes. Mar. Drugs 2009, 7, 210–248. [Google Scholar] [CrossRef] [Green Version]
- Subramani, R.; Aalbersberg, W. Marine actinomycetes: An ongoing source of novel bioactive metabolites. Microbiol. Res. 2012, 167, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.S.; Wellington, E.M.H. The taxonomy of Streptomyces and related genera. Int. J. Syst. Evol. Microbiol. 2001, 51, 797–814. [Google Scholar] [CrossRef] [Green Version]
- Ngamcharungchit, C.; Kanto, H.; Také, A.; Intra, B.; Matsumoto, A.; Panbangred, W.; Inahashi, Y. Amycolatopsis iheyensis sp. nov., isolated from soil on Iheya Island, Japan. Int. J. Syst. Evol. Microbiol. 2023, 73, 005757. [Google Scholar] [CrossRef] [PubMed]
- Citarella, R.V.; Colwell, R.R. Polyphasic Taxonomy of the Genus Vibrio: Polynucleotide Sequence Relationships Among Selected Vibrio Species. J. Bacteriol. 1970, 104, 434. [Google Scholar] [CrossRef]
- Vandamme, P.; Pot, B.; Gillis, M.; De Vos, P.; Kersters, K.; Swings, J. Polyphasic taxonomy, a consensus approach to bacterial systematics. Microbiol. Rev. 1996, 60, 407–438. [Google Scholar] [CrossRef]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Moore, L.H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.; et al. Report of the Ad Hoc Committee on Reconciliation of Approaches to Bacterial Systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Tindall, B.J.; Rosselló-Móra, R.; Busse, H.J.; Ludwig, W.; Kämpfer, P. Notes on the characterization of prokaryote strains for taxonomic purposes. Int. J. Syst. Evol. Microbiol. 2010, 60, 249–266. [Google Scholar] [CrossRef] [Green Version]
- Intra, B.; Matsumoto, A.; Inahashi, Y.; Ōmura, S.; Takahashi, Y.; Panbangred, W. Actinokineospora bangkokensis sp. nov., isolated from rhizospheric soil. Int. J. Syst. Evol. Microbiol. 2013, 63, 2655–2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Intra, B.; Euanorasetr, J.; Také, A.; Inahashi, Y.; Mori, M.; Panbangred, W.; Matsumoto, A. Saccharopolyspora rhizosphaerae sp. nov., an actinomycete isolated from rhizosphere soil in Thailand. Int. J. Syst. Evol. Microbiol. 2019, 69, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Intra, B.; Matsumoto, A.; Inahashi, Y.; Ōmura, S.; Panbangred, W.; Takahashi, Y. Streptosporangium jomthongense sp. nov., an actinomycete isolated from rhizospheric soil and emendation of the genus Streptosporangium. Int. J. Syst. Evol. Microbiol. 2014, 64, 2400–2406. [Google Scholar] [CrossRef] [Green Version]
- Wattanasuepsin, W.; Intra, B.; Také, A.; Inahashi, Y.; Euanorasetr, J.; Ōmura, S.; Matsumoto, A.; Panbangred, W. Saccharomonospora colocasiae sp. Nov., an actinomycete isolated from the rhizosphere of Colocasia esculenta. Int. J. Syst. Evol. Microbiol. 2017, 67, 4572–4577. [Google Scholar] [CrossRef]
- Intra, B.; Panbangred, W.; Inahashi, Y.; Také, A.; Mori, M.; Ōmura, S.; Matsumoto, A. Micromonospora pelagivivens sp. nov., a new species of the genus Micromonospora isolated from deep-sea sediment in Japan. Int. J. Syst. Evol. Microbiol. 2020, 70, 3069–3075. [Google Scholar] [CrossRef]
- Bérdy, J. Thoughts and facts about antibiotics: Where we are now and where we are heading? J. Antibiot. 2012, 65, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Challis, G.L.; Hopwood, D.A. Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species. Proc. Nat. Acad. Sci. USA 2003, 100, 14555–14561. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Kottmann, R.; Yilmaz, P.; Cummings, M.; Biggins, J.B.; Blin, K.; De Bruijn, I.; Chooi, Y.H.; Claesen, J.; Coates, R.C.; et al. Minimum Information about a Biosynthetic Gene cluster. Nat. Chem. Biol. 2015, 11, 625–631. [Google Scholar] [CrossRef]
- Cdc. Antibiotic Resistance Threats Report In The United States 2019. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 16 June 2023).
- De Simeis, D.; Serra, S. Actinomycetes: A Never-Ending Source of Bioactive Compounds—An Overview on Antibiotics Production. Antibiotics 2021, 10, 483. [Google Scholar] [CrossRef]
- Euanorasetr, J.; Nilvongse, A.; Tantimavanich, S.; Nihira, T.; Igarashi, Y.; Panbangred, W. Identification and characterization of soil-isolated Streptomyces SJE177 producing actinomycin. Southeast Asian J. Trop. Med. Public Health 2010, 41, 1177–1187. [Google Scholar]
- Euanorasetr, J.; Intra, B.; Mongkol, P.; Chankhamhaengdecha, S.; Tuchinda, P.; Mori, M.; Shiomi, K.; Nihira, T.; Panbangred, W. Spirotetronate antibiotics with anti-Clostridium activity from Actinomadura sp. 2EPS. W. J. Microbiol. Biotech. 2015, 31, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.; Cruz, J.; Simone, M.; Bernasconi, A.; Brunati, C.; Sosio, M.; Donadio, S.; Maffioli, S.I. Antibacterial Paramagnetic Quinones from Actinoallomurus. J. Nat. Prod. 2017, 80, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Kodani, S.; Komaki, H.; Ishimura, S.; Hemmi, H.; Ohnishi-Kameyama, M. Isolation and structure determination of a new lantibiotic cinnamycin B from Actinomadura atramentaria based on genome mining. J. Ind. Microbiol. Biotechnol. 2016, 43, 1159–1165. [Google Scholar] [CrossRef] [Green Version]
- Shin, B.; Kim, B.Y.; Cho, E.; Oh, K.B.; Shin, J.; Goodfellow, M.; Oh, D.C. Actinomadurol, an Antibacterial Norditerpenoid from a Rare Actinomycete, Actinomadura sp. KC 191. J. Nat. Prod. 2016, 79, 1886–1890. [Google Scholar] [CrossRef] [PubMed]
- Bauermeister, A.; Calil, F.A.; Pinto, F.d.C.L.; Medeiros, T.C.T.; Almeida, L.C.; Silva, L.J.; de Melo, I.S.; Zucchi, T.D.; Costa-Lotufo, L.V.; Moraes, L.A.B. Pradimicin-IRD from Amycolatopsis sp. IRD-009 and its antimicrobial and cytotoxic activities. Nat. Prod. Res. 2019, 33, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Beemelmanns, C.; Ramadhar, T.R.; Kim, K.H.; Klassen, J.L.; Cao, S.; Wyche, T.P.; Hou, Y.; Poulsen, M.; Bugni, T.S.; Currie, C.R.; et al. Macrotermycins A-D, Glycosylated Macrolactams from a Termite-Associated Amycolatopsis sp. M39. Org. Lett. 2017, 19, 1000–1003. [Google Scholar] [CrossRef]
- Khan, A.; Said, M.S.; Borade, B.R.; Gonnade, R.; Barvkar, V.; Kontham, R.; Dastager, S.G. Enceleamycins A-C, Furo-Naphthoquinones from Amycolatopsis sp. MCC0218: Isolation, Structure Elucidation, and Antimicrobial Activity. J. Nat. Prod. 2022, 85, 1267–1273. [Google Scholar] [CrossRef]
- Hashizume, H.; Sawa, R.; Yamashita, K.; Nishimura, Y.; Igarashi, M. Structure and antibacterial activities of new cyclic peptide antibiotics, pargamicins B, C and D, from Amycolatopsis sp. ML1-hF4. J. Ant. 2017, 70, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liang, X.; Zhang, S.; Song, Z.; Wang, C.; Xu, Y. Maipomycin A, a Novel Natural Compound With Promising Anti-biofilm Activity Against Gram-Negative Pathogenic Bacteria. Front. Microbiol. 2021, 11, 598024. [Google Scholar] [CrossRef]
- Kohda, Y.; Sakamoto, S.; Umekita, M.; Kimura, T.; Kubota, Y.; Arisaka, R.; Shibuya, Y.; Muramatsu, H.; Sawa, R.; Dan, S.; et al. Isolation of new derivatives of the 20-membered macrodiolide bispolide from Kitasatospora sp. MG372-hF19. J. Antibiot. 2021, 75, 77–85. [Google Scholar] [CrossRef]
- Uzair, B.; Menaa, F.; Khan, B.A.; Mohammad, F.V.; Ahmad, V.U.; Djeribi, R.; Menaa, B. Isolation, purification, structural elucidation and antimicrobial activities of kocumarin, a novel antibiotic isolated from actinobacterium Kocuria marina CMG S2 associated with the brown seaweed Pelvetia canaliculata. Microbiol. Res. 2018, 206, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Zhen, X.; Li, X.L.; Chen, J.J.; Chen, T.J.; Yang, J.L.; Zhu, P. Tetrocarcin Q, a New Spirotetronate with a Unique Glycosyl Group from a Marine-Derived Actinomycete Micromonospora carbonacea LS276. Mar. Drugs 2018, 16, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, Y.H.; Bae, S.; Sim, J.; Hur, J.; Jo, S.I.; Shin, J.; Suh, Y.G.; Oh, K.B.; Oh, D.C. Nicrophorusamides A and B, Antibacterial Chlorinated Cyclic Peptides from a Gut Bacterium of the Carrion Beetle Nicrophorus concolor. J. Nat. Prod. 2017, 80, 2962–2968. [Google Scholar] [CrossRef] [PubMed]
- Gui, C.; Zhang, S.; Zhu, X.; Ding, W.; Huang, H.; Gu, Y.C.; Duan, Y.; Ju, J. Antimicrobial Spirotetronate Metabolites from Marine-Derived Micromonospora harpali SCSIO GJ089. J. Nat. Prod. 2017, 80, 1594–1603. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xie, L.; Zhao, W.; Zhou, J.; Jiang, H.; Liu, W.; Jiang, H.; Lin, F. Two new rakicidin derivatives from marine Micromonospora chalcea FIM-R160609. Nat. Prod. Res. 2022, 36, 1–8. [Google Scholar] [CrossRef]
- Hifnawy, M.S.; Hassan, H.M.; Mohammed, R.; Fouda, M.M.; Sayed, A.M.; Hamed, A.A.; AbouZid, S.F.; Rateb, M.E.; Alhadrami, H.A.; Abdelmohsen, U.R. Induction of Antibacterial Metabolites by Co-Cultivation of Two Red-Sea-Sponge-Associated Actinomycetes Micromonospora sp. UR56 and Actinokinespora sp. EG49. Mar. Drugs 2020, 18, 243. [Google Scholar] [CrossRef] [PubMed]
- Adnani, N.; Chevrette, M.G.; Adibhatla, S.N.; Zhang, F.; Yu, Q.; Braun, D.R.; Nelson, J.; Simpkins, S.W.; McDonald, B.R.; Myers, C.L.; et al. Coculture of Marine Invertebrate-Associated Bacteria and Interdisciplinary Technologies Enable Biosynthesis and Discovery of a New Antibiotic, Keyicin. ACS Chem. Biol. 2017, 12, 3093–3102. [Google Scholar] [CrossRef]
- Tan, Y.; Hu, Y.; Wang, Q.; Zhou, H.; Wang, Y.; Gan, M. Tetrocarcins N and O, glycosidic spirotetronates from a marine-derived Micromonospora sp. identified by PCR-based screening. RSC Adv. 2016, 6, 91773–91778. [Google Scholar] [CrossRef]
- Pérez-Bonilla, M.; Oves-Costales, D.; De La Cruz, M.; Kokkini, M.; Martín, J.; Vicente, F.; Genilloud, O.; Reyes, F. Phocoenamicins B and C, New Antibacterial Spirotetronates Isolated from a Marine Micromonospora sp. Mar. Drugs 2018, 16, 95. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.E.; Dalisay, D.S.; Chen, J.; Polishchuck, E.A.; Patrick, B.O.; Narula, G.; Ko, M.; Av-Gay, Y.; Li, H.; Magarvey, N.; et al. Aminorifamycins and Sporalactams Produced in Culture by a Micromonospora sp. Isolated from a Northeastern-Pacific Marine Sediment Are Potent Antibiotics. Org. Lett. 2017, 19, 766–769. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhang, Q.; Peng, J.; Zhao, X.; Ma, L.; Zhang, C.; Zhu, Y. Genomics-Driven Discovery of Benzoxazole Alkaloids from the Marine-Derived Micromonospora sp. SCSIO 07395. Molecules 2023, 28, 821. [Google Scholar] [CrossRef]
- Zhou, Q.; Luo, G.C.; Zhang, H.; Tang, G.L. Discovery of 16-Demethylrifamycins by Removing the Predominant Polyketide Biosynthesis Pathway in Micromonospora sp. Strain TP-A0468. Appl. Environ. Microbiol. 2019, 85, e02597-18. [Google Scholar] [CrossRef] [Green Version]
- Lasch, C.; Gummerlich, N.; Myronovskyi, M.; Palusczak, A.; Zapp, J.; Luzhetskyy, A. Loseolamycins: A Group of New Bioactive Alkylresorcinols Produced after Heterologous Expression of a Type III PKS from Micromonospora endolithica. Molecules 2020, 25, 4594. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wen, Z.; Liu, L.; Zhu, X.; Shen, B.; Yan, X.; Duan, Y.; Huang, Y. Yangpumicins F and G, Enediyne Congeners from Micromonospora yangpuensis DSM 45577. J. Nat. Prod. 2019, 82, 2483–2488. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Li, Y.-J.; Wang, Z.-M.; Wang, Y.-F.; Wang, B.; Shi, D.-Y. New Pyrroline Isolated from Antarctic Krill-Derived Actinomycetes Nocardiopsis sp. LX-1 Combining with Molecular Networking. Mar. Drugs 2023, 21, 127. [Google Scholar] [CrossRef] [PubMed]
- Siddharth, S.; Aswathanarayan, J.B.; Kuruburu, M.G.; Madhunapantula, S.R.V.; Vittal, R.R. Diketopiperazine derivative from marine actinomycetes Nocardiopsis sp. SCA30 with antimicrobial activity against MRSA. Arch. Microbiol. 2021, 203, 6173–6181. [Google Scholar] [CrossRef]
- Braña, A.F.; Sarmiento-Vizcaíno, A.; Pérez-Victoria, I.; Otero, L.; Fernández, J.; Palacios, J.J.; Martín, J.; De La Cruz, M.; Díaz, C.; Vicente, F.; et al. Branimycins B and C, Antibiotics Produced by the Abyssal Actinobacterium Pseudonocardia carboxydivorans M-227. J. Nat. Prod. 2017, 80, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Matroodi, S.; Siitonen, V.; Baral, B.; Yamada, K.; Akhgari, A.; Metsä-Ketelä, M. Genotyping-Guided Discovery of Persiamycin A From Sponge-Associated Halophilic Streptomonospora sp. PA3. Front Microbiol. 2020, 11, 1237. [Google Scholar] [CrossRef]
- Pereira, F.; Almeida, J.R.; Paulino, M.; Grilo, I.R.; Macedo, H.; Cunha, I.; Sobral, R.G.; Vasconcelos, V.; Gaudêncio, S.P. Antifouling Napyradiomycins from Marine-Derived Actinomycetes Streptomyces aculeolatus. Mar. Drugs 2020, 18, 63. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Zhang, C.; Liu, Y.; Huang, Y.; Bai, Y.; Hang, X.; Zeng, L.; Zhu, D.; Bi, H. Armeniaspirol A: A novel anti-Helicobacter pylori agent. Microb. Biotechnol. 2022, 15, 442–454. [Google Scholar] [CrossRef]
- Rodríguez Estévez, M.; Gummerlich, N.; Myronovskyi, M.; Zapp, J.; Luzhetskyy, A. Benzanthric Acid, a Novel Metabolite From Streptomyces albus Del14 Expressing the Nybomycin Gene Cluster. Front. Chem. 2020, 7, 896. [Google Scholar] [CrossRef] [PubMed]
- Manikkam, R.; Murthy, S.; Palaniappan, S.; Kaari, M.; Sahu, A.K.; Said, M.; Ganesan, V.; Kannan, S.; Ramasamy, B.; Thirugnanasambandan, S. Antibacterial and Anti-HIV Metabolites from Marine Streptomyces albus MAB56 Isolated from Andaman and Nicobar Islands, India. Appl. Biochem. Biotechnol. 2023, 195, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.; Kwon, O.S.; Shin, J.; Oh, K.B. Antibacterial Activity and Mode of Action of Lactoquinomycin A from Streptomyces bacillaris. Mar. Drugs 2020, 19, 7. [Google Scholar] [CrossRef]
- Singh, R.; Dubey, A.K. Isolation and Characterization of a New Endophytic Actinobacterium Streptomyces californicus Strain ADR1 as a Promising Source of Anti-Bacterial, Anti-Biofilm and Antioxidant Metabolites. Microorganisms 2020, 8, 929. [Google Scholar] [CrossRef] [PubMed]
- Shaala, L.A.; Youssef, D.T.A.; Alzughaibi, T.A.; Elhady, S.S. Antimicrobial Chlorinated 3-Phenylpropanoic Acid Derivatives from the Red Sea Marine Actinomycete Streptomyces coelicolor LY001. Mar. Drugs 2020, 18, 450. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Chen, N.; Li, J.; Su, J.C.; Yang, J.; Zhang, C.X.; Lin, H.W.; Zhou, Y. Antimicrobial Chlorinated Carbazole Alkaloids from the Sponge-Associated Actinomycete Streptomyces diacarni LHW51701. Chin. J. Chem. 2021, 39, 1188–1192. [Google Scholar] [CrossRef]
- Takehana, Y.; Umekita, M.; Hatano, M.; Kato, C.; Sawa, R.; Igarashi, M. Fradiamine A, a new siderophore from the deep-sea actinomycete Streptomyces fradiae MM456M-mF7. J. Antibiot. 2017, 70, 611–615. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Ou, P.; Liu, L.; Jin, X. Anti-MRSA Activity of Actinomycin X2 and Collismycin A Produced by Streptomyces globisporus WA5-2-37 From the Intestinal Tract of American Cockroach (Periplaneta americana). Front. Microbiol. 2020, 11, 555. [Google Scholar] [CrossRef] [Green Version]
- Kaweewan, I.; Komaki, H.; Hemmi, H.; Kodani, S. Isolation and structure determination of a new thiopeptide globimycin from Streptomyces globisporus subsp. globisporus based on genome mining. Tetrahedron Lett. 2018, 59, 409–414. [Google Scholar] [CrossRef]
- Saleem, M.; Hassan, A.; Li, F.; Lu, Q.; Ponomareva, L.V.; Parkin, S.; Sun, C.; Thorson, J.S.; Shaaban, K.A.; Sajid, I. Bioprospecting of desert actinobacteria with special emphases on griseoviridin, mitomycin C and a new bacterial metabolite producing Streptomyces sp. PU-KB10–4. BMC Microbiol. 2023, 23, 69. [Google Scholar] [CrossRef]
- Harunari, E.; Imada, C.; Igarashi, Y. Konamycins A and B and Rubromycins CA1 and CA2, Aromatic Polyketides from the Tunicate-Derived Streptomyces hyaluromycini MB-PO13T. J. Nat. Prod. 2019, 82, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Song, Y.; Li, X.; Wang, X.; Ling, C.; Qin, X.; Zhou, Z.; Li, Q.; Wei, X.; Ju, J. Abyssomicin Monomers and Dimers from the Marine-Derived Streptomyces koyangensis SCSIO 5802. J. Nat. Prod. 2018, 81, 1892–1898. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.; Kwon, O.S.; Chung, B.; Lee, J.; Sun, J.; Shin, J.; Oh, K.B. Antibacterial Activity of Chromomycins from a Marine-Derived Streptomyces microflavus. Mar. Drugs 2020, 18, 522. [Google Scholar] [CrossRef]
- Martinet, L.; Naômé, A.; Rezende, L.C.; Tellatin, D.; Pignon, B.; Docquier, J.-D.; Sannio, F.; Baiwir, D.; Mazzucchelli, G.; Frédérich, M. Lunaemycins, new cyclic hexapeptide antibiotics from the cave moonmilk-dweller Streptomyces lunaelactis MM109T. Int. J. Mol. Sci. 2023, 24, 1114. [Google Scholar] [CrossRef]
- Yang, L.; Hou, L.; Li, H.; Li, W. Antibiotic angucycline derivatives from the deepsea-derived Streptomyces lusitanus. Nat. Prod. Res. 2020, 34, 3444–3450. [Google Scholar] [CrossRef] [PubMed]
- Sujarit, K.; Mori, M.; Dobashi, K.; Shiomi, K.; Pathom-Aree, W.; Lumyong, S. New Antimicrobial Phenyl Alkenoic Acids Isolated from an Oil Palm Rhizosphere-Associated Actinomycete, Streptomyces palmae CMU-AB204T. Microorganisms 2020, 8, 350. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Tian, E.; Kong, F.; Hong, K. Bioactive Molecules from Mangrove Streptomyces qinglanensis 172205. Mar. Drugs 2020, 18, 255. [Google Scholar] [CrossRef]
- Heo, C.-S.; Kang, J.S.; Kwon, J.-H.; Anh, C.V.; Shin, H.J. Pyrrole-Containing Alkaloids from a Marine-Derived Actinobacterium Streptomyces zhaozhouensis and Their Antimicrobial and Cytotoxic Activities. Mar. Drugs 2023, 21, 167. [Google Scholar] [CrossRef] [PubMed]
- Bo, S.T.; Xu, Z.F.; Yang, L.; Cheng, P.; Tan, R.X.; Jiao, R.H.; Ge, H.M. Structure and biosynthesis of mayamycin B, a new polyketide with antibacterial activity from Streptomyces sp. 120454. J. Antibiot. 2018, 71, 601–605. [Google Scholar] [CrossRef]
- Liang, Y.; Xie, X.; Chen, L.; Yan, S.; Ye, X.; Anjum, K.; Huang, H.; Lian, X.; Zhang, Z. Bioactive Polycyclic Quinones from Marine Streptomyces sp. 182SMLY. Mar. Drugs 2016, 14, 10. [Google Scholar] [CrossRef] [Green Version]
- Safaei, N.; Mast, Y.; Steinert, M.; Huber, K.; Bunk, B.; Wink, J. Angucycline-like Aromatic Polyketide from a Novel Streptomyces Species Reveals Freshwater Snail Physa acuta as Underexplored Reservoir for Antibiotic-Producing Actinomycetes. Antibiotics 2020, 10, 22. [Google Scholar] [CrossRef]
- Wang, T.; Li, F.; Lu, Q.; Wu, G.; Jiang, Z.; Liu, S.; Habden, X.; Razumova, E.A.; Osterman, I.A.; Sergiev, P.V.; et al. Diversity, novelty, antimicrobial activity, and new antibiotics of cultivable endophytic actinobacteria isolated from psammophytes collected from Taklamakan Desert. J. Pharm. Anal. 2021, 11, 241–250. [Google Scholar] [CrossRef]
- Guerrero-Garzón, J.F.; Zehl, M.; Schneider, O.; Rückert, C.; Busche, T.; Kalinowski, J.; Bredholt, H.; Zotchev, S.B. Streptomyces spp. From the Marine Sponge Antho dichotoma: Analyses of Secondary Metabolite Biosynthesis Gene Clusters and Some of Their Products. Front. Microbiol. 2020, 11, 437. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Wang, X.; Huang, T.; Deng, Z.; Lin, S. Naphthoquinone-Based Meroterpenoids from Marine-Derived Streptomyces sp. B9173. Biomolecules 2020, 10, 1187. [Google Scholar] [CrossRef]
- Carretero-Molina, D.; Ortiz-López, F.J.; Martín, J.; Oves-Costales, D.; Díaz, C.; De La Cruz, M.; Cautain, B.; Vicente, F.; Genilloud, O.; Reyes, F. New Napyradiomycin Analogues from Streptomyces sp. Strain CA-271078. Mar. Drugs 2019, 18, 22. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Hu, X.; Hu, X.; Li, S.; Li, L.; Yu, L.; Liu, H.; You, X.; Wang, Z.; Li, L.; et al. Cytotoxic and Antibacterial Cervinomycins B1-4 from a Streptomyces Species. J. Nat. Prod. 2019, 82, 2337–2342. [Google Scholar] [CrossRef] [PubMed]
- Dardić, D.; Lauro, G.; Bifulco, G.; Laboudie, P.; Sakhaii, P.; Bauer, A.; Vilcinskas, A.; Hammann, P.E.; Plaza, A. Svetamycins A-G, Unusual Piperazic Acid-Containing Peptides from Streptomyces sp. J. Org. Chem. 2017, 82, 6032–6043. [Google Scholar] [CrossRef] [PubMed]
- Lü, Y.; Shao, M.; Wang, Y.; Qian, S.; Wang, M.; Wang, Y.; Li, X.; Bao, Y.; Deng, C.; Yue, C.; et al. Zunyimycins B and C, New Chloroanthrabenzoxocinones Antibiotics against Methicillin-Resistant Staphylococcus aureus and Enterococci from Streptomyces sp. FJS31-2. Molecules 2017, 22, 251. [Google Scholar] [CrossRef] [Green Version]
- Kitani, S.; Ueguchi, T.; Igarashi, Y.; Leetanasaksakul, K.; Thamchaipenet, A.; Nihira, T. Rakicidin F, a new antibacterial cyclic depsipeptide from a marine sponge-derived Streptomyces sp. J. Ant. 2017, 71, 139–141. [Google Scholar] [CrossRef]
- Yang, Z.; Shao, L.; Wang, M.; Rao, M.; Ge, M.; Xu, Y. Two novel quinomycins discovered by UPLC-MS from Stretomyces sp. HCCB11876. J. Ant. 2019, 72, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.X.; Ding, R.; Jiang, S.T.; Tang, J.S.; Hu, D.; Chen, G.D.; Lin, F.; Hong, K.; Yao, X.S.; Gao, H. Aldgamycins J-O, 16-Membered Macrolides with a Branched Octose Unit from Streptomycetes sp. and Their Antibacterial Activities. J. Nat. Prod. 2016, 79, 2446–2454. [Google Scholar] [CrossRef]
- Zhou, B.; Ji, Y.Y.; Zhang, H.J.; Shen, L. Gephyyamycin and cysrabelomycin, two new angucyclinone derivatives from the Streptomyces sp. HN-A124. Nat. Prod. Res. 2021, 35, 2117–2122. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qi, H.; Zhang, H.; Zhang, S.-Y.; Zhang, C.-H.; Zhang, L.-Q.; Xiang, W.-S.; Wang, J.-D. Anulamycins A–F, Cinnamoyl-Containing Peptides from a Lake Sediment Derived Streptomyces. J. Nat. Prod. 2023, 86, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, L.; Zhang, X.; Liang, Y.; Anjum, K.; Chen, L.; Lian, X.Y. Bioactive Bafilomycins and a New N-Arylpyrazinone Derivative from Marine-derived Streptomyces sp. HZP-2216E. Planta Med. 2017, 83, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Voitsekhovskaia, I.; Paulus, C.; Dahlem, C.; Rebets, Y.; Nadmid, S.; Zapp, J.; Axenov-Gribanov, D.; Rückert, C.; Timofeyev, M.; Kalinowski, J.; et al. Baikalomycins A-C, New Aquayamycin-Type Angucyclines Isolated from Lake Baikal Derived Streptomyces sp. IB201691-2A. Microorganisms 2020, 8, 680. [Google Scholar] [CrossRef]
- Iniyan, A.M.; Sudarman, E.; Wink, J.; Kannan, R.R.; Vincent, S.G.P. Ala-geninthiocin, a new broad spectrum thiopeptide antibiotic, produced by a marine Streptomyces sp. ICN19. J. Antibiot. 2019, 72, 99–105. [Google Scholar] [CrossRef]
- Son, S.; Hong, Y.S.; Jang, M.; Heo, K.T.; Lee, B.; Jang, J.P.; Kim, J.W.; Ryoo, I.J.; Kim, W.G.; Ko, S.K.; et al. Genomics-Driven Discovery of Chlorinated Cyclic Hexapeptides Ulleungmycins A and B from a Streptomyces Species. J. Nat. Prod. 2017, 80, 3025–3031. [Google Scholar] [CrossRef]
- Son, S.; Ko, S.K.; Kim, S.M.; Kim, E.; Kim, G.S.; Lee, B.; Ryoo, I.J.; Kim, W.G.; Lee, J.S.; Hong, Y.S.; et al. Antibacterial Cyclic Lipopeptide Enamidonins with an Enamide-Linked Acyl Chain from a Streptomyces Species. J. Nat. Prod. 2018, 81, 2462–2469. [Google Scholar] [CrossRef]
- Sawa, R.; Kubota, Y.; Umekita, M.; Hatano, M.; Hayashi, C.; Igarashi, M. Quadoctomycin, a 48-membered macrolide antibiotic from Streptomyces sp. MM168-141F8. J. Antibiot. 2017, 71, 91–96. [Google Scholar] [CrossRef]
- Konwar, A.N.; Basak, S.; Devi, S.G.; Borah, J.C.; Thakur, D. Streptomyces sp. MNP32, a forest-dwelling Actinomycetia endowed with potent antibacterial metabolites. 3 Biotech 2023, 13, 257. [Google Scholar] [CrossRef]
- Zhang, L.; Shi, J.; Liu, C.L.; Xiang, L.; Ma, S.Y.; Li, W.; Jiao, R.H.; Tan, R.X.; Ge, H.M. New borrelidin derivatives from an endophytic Streptomyces sp. Tetrahedron Lett. 2018, 59, 4517–4520. [Google Scholar] [CrossRef]
- Guo, Z.K.; Wang, Y.C.; Tan, Y.Z.; Abulaizi, A.; Xiong, Z.J.; Zhang, S.Q.; Yang, Y.; Yang, L.Y.; Shi, J. Nagimycins A and B, Antibacterial Ansamycin-Related Macrolactams from Streptomyces sp. NA07423. Org. Lett. 2023, 25, 4203–4207. [Google Scholar] [CrossRef] [PubMed]
- Arn, F.; Frasson, D.; Kroslakova, I.; Rezzonico, F.; Pothie, J.F.; Riedl, R.; Sievers, M. Isolation and Identification of Actinomycetes Strains from Switzerland and their Biotechnological Potential. Chimia 2020, 74, 382–390. [Google Scholar] [CrossRef]
- Mazumdar, R.; Dutta, P.P.; Saikia, J.; Borah, J.C.; Thakur, D. Streptomyces sp. Strain PBR11, a Forest-Derived Soil Actinomycetia with Antimicrobial Potential. Microbiol. Spectr. 2023, 11, 17544. [Google Scholar] [CrossRef]
- Betancur, L.A.; Forero, A.M.; Vinchira-Villarraga, D.M.; Cárdenas, J.D.; Romero-Otero, A.; Chagas, F.O.; Pupo, M.T.; Castellanos, L.; Ramos, F.A. NMR-based metabolic profiling to follow the production of anti-phytopathogenic compounds in the culture of the marine strain Streptomyces sp. PNM-9. Microbiol. Res. 2020, 239, 126507. [Google Scholar] [CrossRef]
- Cheng, C.; Othman, E.M.; Reimer, A.; Grüne, M.; Kozjak-Pavlovic, V.; Stopper, H.; Hentschel, U.; Abdelmohsen, U.R. Ageloline A, new antioxidant and antichlamydial quinolone from the marine sponge-derived bacterium Streptomyces sp. SBT345. Tetrahedron Lett. 2016, 57, 2786–2789. [Google Scholar] [CrossRef]
- Cong, Z.; Huang, X.; Liu, Y.; Liu, Y.; Wang, P.; Liao, S.; Yang, B.; Zhou, X.; Huang, D.; Wang, J. Cytotoxic anthracycline and antibacterial tirandamycin analogues from a marine-derived Streptomyces sp. SCSIO 41399. J. Antibiot. 2019, 72, 45–49. [Google Scholar] [CrossRef]
- Tian, H.; Shafi, J.; Ji, M.; Bi, Y.; Yu, Z. Antimicrobial Metabolites from Streptomyces sp. SN0280. J. Nat. Prod. 2017, 80, 1015–1019. [Google Scholar] [CrossRef]
- Maiti, P.K.; Das, S.; Sahoo, P.; Mandal, S. Streptomyces sp. SM01 isolated from Indian soil produces a novel antibiotic picolinamycin effective against multi drug resistant bacterial strains. Sci. Rep. 2020, 10, 788. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, X.; He, N.; Xie, Y.; Hong, B. Rescrutiny of the sansanmycin biosynthetic gene cluster leads to the discovery of a novel sansanmycin analogue with more potency against Mycobacterium tuberculosis. J. Antibiot. 2019, 72, 769–774. [Google Scholar] [CrossRef]
- Pratiwi, R.H.; Hidayat, I.; Hanafi, M.; Mangunwardoyo, W. Isolation and structure elucidation of phenazine derivative from Streptomyces sp. strain UICC B-92 isolated from Neesia altissima (Malvaceae). Iran. J. Microbiol. 2020, 12, 127. [Google Scholar] [CrossRef]
- Jiang, Y.J.; Zhang, D.S.; Zhang, H.J.; Li, J.Q.; Ding, W.J.; Xu, C.D.; Ma, Z.J. Medermycin-Type Naphthoquinones from the Marine-Derived Streptomyces sp. XMA39. J. Nat. Prod. 2018, 81, 2120–2124. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ren, Z.; Chunyu, W.X.; Li, G.D.; Chen, X.; Zhang, Z.T.L.; Sun, H.B.; Wang, M.; Xie, T.P.; Wang, M.; et al. Exploration of Diverse Secondary Metabolites From Streptomyces sp. YINM00001, Using Genome Mining and One Strain Many Compounds Approach. Front. Microbiol. 2022, 13, 831174. [Google Scholar] [CrossRef] [PubMed]
- Newaz, A.W.; Yong, K.; Lian, X.Y.; Zhang, Z. Streptoindoles A–D, novel antimicrobial indole alkaloids from the marine-associated actinomycete Streptomyces sp. ZZ1118. Tetrahedron 2022, 104, 132598. [Google Scholar] [CrossRef]
- Zhang, D.; Yi, W.; Ge, H.; Zhang, Z.; Wu, B. Bioactive Streptoglutarimides A-J from the Marine-Derived Streptomyces sp. ZZ741. J. Nat. Prod. 2019, 82, 2800–2808. [Google Scholar] [CrossRef] [PubMed]
- Asolkar, R.N.; Singh, A.; Jensen, P.R.; Aalbersberg, W.; Carté, B.K.; Feussner, K.D.; Subramani, R.; DiPasquale, A.; Rheingold, A.L.; Fenical, W. Marinocyanins, cytotoxic bromo-phenazinone meroterpenoids from a marine bacterium from the streptomycete clade MAR4. Tetrahedron 2017, 73, 2234–2241. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y.; Kagoshima, Y.; Masuda, T.; Kizuka, M.; Ogawa, Y.; Endo, S.; Nishigoori, H.; Saito, K.; Takasugi, K.; Miura, M.; et al. Muraminomicins, new lipo-nucleoside antibiotics from Streptosporangium sp. SANK 60501-structure elucidations of muraminomicins and supply of the core component for derivatization. J. Antibiot. 2019, 72, 943–955. [Google Scholar] [CrossRef]
- Teta, R.; Marteinsson, V.T.; Longeon, A.; Klonowski, A.M.; Groben, R.; Bourguet-Kondracki, M.L.; Costantino, V.; Mangoni, A. Thermoactinoamide A, an Antibiotic Lipophilic Cyclopeptide from the Icelandic Thermophilic Bacterium Thermoactinomyces vulgaris. J. Nat. Prod. 2017, 80, 2530–2535. [Google Scholar] [CrossRef]
- Zhang, S.; Xie, Q.; Sun, C.; Tian, X.P.; Gui, C.; Qin, X.; Zhang, H.; Ju, J. Cytotoxic Kendomycins Containing the Carbacylic Ansa Scaffold from the Marine-Derived Verrucosispora sp. SCSIO 07399. J. Nat. Prod. 2019, 82, 3366–3371. [Google Scholar] [CrossRef]
- Gupte, M.; Kulkarni, P.; Ganguli, B.N. Antifungal antibiotics. Appl. Microbiol. Biotechnol. 2002, 58, 46–57. [Google Scholar] [CrossRef]
- Hazen, E.L.; Brown, R. Fungicidin, an Antibiotic Produced by a Soil Actinomycete. Proc. Soc. Exp. Biol. Med. 1951, 76, 93–97. [Google Scholar] [CrossRef]
- Georgopapadakou, N.H. Antifungals: Mechanism of action and resistance, established and novel drugs. Curr. Opin. Microbiol. 1998, 1, 547–557. [Google Scholar] [CrossRef]
- Ghannoum, M.A.; Rice, L.B. Antifungal agents: Mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Intra, B.; Euanorasetr, J.; Nihira, T.; Panbangred, W. Characterization of a gamma-butyrolactone synthetase gene homologue (stcA) involved in bafilomycin production and aerial mycelium formation in Streptomyces sp. SBI034. Appl. Microbiol. Biotechnol. 2016, 100, 2749–2760. [Google Scholar] [CrossRef] [PubMed]
- Intra, B.; Greule, A.; Bechthold, A.; Euanorasetr, J.; Paululat, T.; Panbangred, W. Thailandins A and B, New Polyene Macrolactone Compounds Isolated from Actinokineospora bangkokensis Strain 44EHWT, Possessing Antifungal Activity against Anthracnose Fungi and Pathogenic Yeasts. J. Agric. Food Chem. 2016, 64, 5171–5179. [Google Scholar] [CrossRef] [PubMed]
- Euanorasetr, J.; Junhom, M.; Tantimavanich, S.; Vorasin, O.; Munyoo, B.; Tuchinda, P.; Panbangred, W. Halogenated benzoate derivatives of altholactone with improved anti-fungal activity. J. Asian Nat. Prod. Res. 2016, 18, 462–474. [Google Scholar] [CrossRef]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Suriyachadkun, C.; Supothina, S.; Chanthaket, R.; Chutrakul, C.; Vichai, V. Actinomadurone, a polycyclic tetrahydroxanthone from Actinomadura sp. BCC 35430. Tetrahedron Lett. 2017, 58, 3223–3225. [Google Scholar] [CrossRef]
- Cheng, M.-J.; Chen, J.-J.; Wu, M.-D.; Leu, J.-Y.; Tseng, M. Antifungal Activities of Compounds Produced by Newly Isolated Acrocarpospora Strains. Antibiotics 2023, 12, 95. [Google Scholar] [CrossRef]
- Kim, H.R.; Kim, J.; Yu, J.S.; Lee, B.S.; Kim, K.H.; Kim, C.S. Isolation, structure elucidation, total synthesis, and biosynthesis of dermazolium A, an antibacterial imidazolium metabolite of a vaginal bacterium Dermabacter vaginalis. Arch. Pharm. Res. 2023, 46, 35–43. [Google Scholar] [CrossRef]
- Hara, S.; Ishikawa, N.; Hara, Y.; Nehira, T.; Sakai, K.; Gonoi, T.; Ishibashi, M. Nabscessins A and B, Aminocyclitol Derivatives from Nocardia abscessus IFM 10029T. J. Nat. Prod. 2017, 80, 565–568. [Google Scholar] [CrossRef]
- Kim, H.J.; Han, C.Y.; Park, J.S.; Oh, S.H.; Kang, S.H.; Choi, S.S.; Kim, J.M.; Kwak, J.H.; Kim, E.S. Nystatin-like Pseudonocardia polyene B1, a novel disaccharide-containing antifungal heptaene antibiotic. Sci. Rep. 2018, 8, 13584. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Gao, X.; Kang, Z.; Huang, L.; Fan, D.; Yan, X. Saccharothrix yanglingensis Strain Hhs.015 Is a Promising Biocontrol Agent on Apple Valsa Canker. Plant Dis. 2016, 100, 510–514. [Google Scholar] [CrossRef] [Green Version]
- Thekkangil, A.; George, B.; Prakash, S.M.U.; Suchithra, T.V. Mechanism of Streptomyces albidoflavus STV1572a derived 1-heneicosanol as an inhibitor against squalene epoxidase of Trichophyton mentagrophytes. Microb. Pathog. 2021, 154, 104853. [Google Scholar] [CrossRef]
- Ding, N.; Jiang, Y.; Han, L.; Chen, X.; Ma, J.; Qu, X.; Mu, Y.; Liu, J.; Li, L.; Jiang, C.; et al. Bafilomycins and Odoriferous Sesquiterpenoids from Streptomyces albolongus Isolated from Elephas maximus Feces. J. Nat. Prod. 2016, 79, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Fu, S.N.; Bao, Y.X.; Yang, Y.; Shen, H.F.; Lin, B.R.; Zhou, G.X. Kitamycin C, a new antimycin-type antibiotic from Streptomyces antibioticus strain 200-09. Nat. Prod. Res. 2017, 31, 1819–1824. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.; Schleissner, C.; Fernández, R.; Rodríguez, P.; Reyes, F.; Zuñiga, P.; De La Calle, F.; Cuevas, C. PM100117 and PM100118, new antitumor macrolides produced by a marine Streptomyces caniferus GUA-06-05-006A. J. Antibiot. 2016, 69, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Vicente Dos Reis, G.; Abraham, W.R.; Grigoletto, D.F.; De Campos, J.B.; Marcon, J.; Da Silva, J.A.; Quecine, M.C.; De Azevedo, J.L.; Ferreira, A.G.; De Lira, S.P. Gloeosporiocide, a new antifungal cyclic peptide from Streptomyces morookaense AM25 isolated from the Amazon bulk soil. FEMS Microbiol. Lett. 2019, 366, 175. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, Z.; Zhao, J.; Zhuang, X.; Cao, P.; Guo, X.; Liu, C.; Xiang, W. Community Composition, Antifungal Activity and Chemical Analyses of Ant-Derived Actinobacteria. Front. Microbiol. 2020, 11, 201. [Google Scholar] [CrossRef] [Green Version]
- Mojicevic, M.; D’Agostino, P.M.; Pavic, A.; Vojnovic, S.; Senthamaraikannan, R.; Vasiljevic, B.; Gulder, T.A.M.; Nikodinovic-Runic, J. Streptomyces sp. BV410 isolate from chamomile rhizosphere soil efficiently produces staurosporine with antifungal and antiangiogenic properties. Microbiol. Open 2020, 9, e986. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Peng, D.; Peng, F.-F.; Zhang, Q.-B.; Duan, Y.-W.; Huang, Y. The Isolation and Structure Elucidation of Spirotetronate Lobophorins A, B, and H8 from Streptomyces sp. CB09030 and Their Biosynthetic Gene Cluster. Molecules 2023, 28, 3597. [Google Scholar] [CrossRef]
- Fang, Q.; Maglangit, F.; Mugat, M.; Urwald, C.; Kyeremeh, K.; Deng, H. Targeted Isolation of Indole Alkaloids from Streptomyces sp. CT37. Molecules 2020, 25, 1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Niu, H.J.; Qu, T.L.; Zhang, X.F.; Du, F.Y. Streptomyces sp. FX13 inhibits fungicide-resistant Botrytis cinerea in vitro and in vivo by producing oligomycin A. Pestic. Biochem. Physiol. 2021, 175, 104834. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Li, C.; Wang, H.; Yu, Z.; Xu, X.; Wang, X.; Zhao, J.; Xiang, W. Community Structures and Antifungal Activity of Root-Associated Endophytic Actinobacteria in Healthy and Diseased Cucumber Plants and Streptomyces sp. HAAG3-15 as a Promising Biocontrol Agent. Microorganisms 2020, 8, 236. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Wang, L.; Yang, J.; Zhang, F.; Sun, Y.; Yu, M.; Yan, Y.; Ma, Y.T.; Huang, S.X. A new antifungal macrolide from Streptomyces sp. KIB-H869 and structure revision of halichomycin. Tetrahedron Lett. 2016, 57, 1375–1378. [Google Scholar] [CrossRef]
- Feng, X.-L.; Zhang, R.-Q.; Wang, D.-C.; Dong, W.-G.; Wang, Z.-X.; Zhai, Y.-J.; Han, W.-B.; Yin, X.; Tian, J.; Wei, J. Genomic and Metabolite Profiling Reveal a Novel Streptomyces Strain, QHH-9511, from the Qinghai-Tibet Plateau. Microbiol. Spectr. 2023, 11, e02764-02722. [Google Scholar] [CrossRef] [PubMed]
- Yi, W.; Qin, L.; Lian, X.Y.; Zhang, Z. New Antifungal Metabolites from the Mariana Trench Sediment-Associated Actinomycete Streptomyces sp. SY1965. Mar. Drugs 2020, 18, 385. [Google Scholar] [CrossRef]
- Herbrík, A.; Corretto, E.; Chroňáková, A.; Langhansová, H.; Petrásková, P.; Hrdý, J.; Čihák, M.; Krištůfek, V.; Bobek, J.; Petříček, M.; et al. A Human Lung-Associated Streptomyces sp. TR1341 Produces Various Secondary Metabolites Responsible for Virulence, Cytotoxicity and Modulation of Immune Response. Front. Microbiol. 2020, 10, 3028. [Google Scholar] [CrossRef] [Green Version]
- Dutta, J.; Thakur, D. Evaluation of Antagonistic and Plant Growth Promoting Potential of Streptomyces sp. TT3 Isolated from Tea (Camellia sinensis) Rhizosphere Soil. Curr. Microbiol. 2020, 77, 1829–1838. [Google Scholar] [CrossRef]
- Yamamoto, K.; Futamura, Y.; Uson-Lopez, R.A.; Aono, H.; Shimizu, T.; Osada, H. YO-001A, a new antifungal agent produced by Streptomyces sp. YO15-A001. J. Antibiot. 2019, 72, 986–990. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Sharma, R.; Srinivas, V.; Naresh, N.; Mishra, S.P.; Ankati, S.; Pratyusha, S.; Govindaraj, M.; Gonzalez, S.V.; Nervik, S.; et al. Identification and Characterization of a Streptomyces albus Strain and Its Secondary Metabolite Organophosphate against Charcoal Rot of Sorghum. Plants 2020, 9, 1727. [Google Scholar] [CrossRef]
- Hoshino, S.; Wong, C.P.; Ozeki, M.; Zhang, H.; Hayashi, F.; Awakawa, T.; Asamizu, S.; Onaka, H.; Abe, I. Umezawamides, new bioactive polycyclic tetramate macrolactams isolated from a combined-culture of Umezawaea sp. and mycolic acid-containing bacterium. J. Antibiot. 2018, 71, 653–657. [Google Scholar] [CrossRef]
- Barreiro, C.; Prieto, C.; Sola-Landa, A.; Solera, E.; Martínez-Castro, M.; Pérez-Redondo, R.; García-Estrada, C.; Aparicio, J.F.; Fernández-Martínez, L.T.; Santos-Aberturas, J.; et al. Draft genome of Streptomyces tsukubaensis NRRL 18488, the producer of the clinically important immunosuppressant tacrolimus (FK506). J. Bacteriol. 2012, 194, 3756–3757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bierer, B.E.; Mattila, P.S.; Standaert, R.F.; Herzenberg, L.A.; Burakoff, S.J.; Crabtree, G.; Schreiber, S.L. Two distinct signal transmission pathways in T lymphocytes are inhibited by complexes formed between an immunophilin and either FK506 or rapamycin. Proc. Nat. Acad. Sci. USA 1990, 87, 9231–9235. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Ikeda, M. Deodorization of pig feces by actinomycetes. Appl. Environ. Microbiol. 1978, 36, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Murata, A.; Shinsaku, H. Accelerated composting of cereal shochu-distillery wastes by actinomycetes. J. Ferment. Bioeng. 1995, 80, 421. [Google Scholar] [CrossRef]
- Mansour, F.A.; Mohamedin, A.H. Enzymes of Candida albicans cell-wall lytic system produced by Streptomyces thermodiastaticus. Acta Microbiol. Immunol. Hung. 2001, 48, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant Growth Promoting and Biocontrol Activity of Streptomyces spp. as Endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [Green Version]
- Subramani, R.; Sipkema, D. Marine Rare Actinomycetes: A Promising Source of Structurally Diverse and Unique Novel Natural Products. Mar. Drugs 2019, 17, 249. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Jeong, G.S. Salinosporamide A, a Marine-Derived Proteasome Inhibitor, Inhibits T Cell Activation through Regulating Proliferation and the Cell Cycle. Molecules 2020, 25, 5031. [Google Scholar] [CrossRef]
- Arcamone, F.; Cassinelli, G.; Fantini, G.; Grein, A.; Orezzi, P.; Pol, C.; Spalla, C. Adriamycin, 14-hydroxydaimomycin, a new antitumor antibiotic from S. peucetius var. caesius. Biotechnol. Bioeng. 1969, 11, 1101–1110. [Google Scholar] [CrossRef]
- Jagannathan, S.V.; Manemann, E.M.; Rowe, S.E.; Callender, M.C.; Soto, W. Marine Actinomycetes, New Sources of Biotechnological Products. Mar. Drugs 2021, 19, 365. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, Y.; Shimasaki, R.; Miyanaga, S.; Oku, N.; Onaka, H.; Sakurai, H.; Saiki, I.; Kitani, S.; Nihira, T.; Wimonsiravude, W.; et al. Rakicidin D, an inhibitor of tumor cell invasion from marine-derived Streptomyces sp. J. Antibiot. 2010, 63, 563–565. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Chen, Y.; Ukaji, T.; Okada, S.; Umezawa, K. Isolation of ketomycin from Actinomycetes as an inhibitor of 2D and 3D cancer cell invasion. J. Antibiot. 2018, 72, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.K.; Lee, H.S.; Kang, J.S.; Shin, H.J. Dokdolipids A−C, Hydroxylated Rhamnolipids from the Marine-Derived Actinomycete Actinoalloteichus hymeniacidonis. Mar. Drugs 2019, 17, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, T.; Iwatsuki, M.; Asami, Y.; Ishiyama, A.; Hokari, R.; Otoguro, K.; Matsumoto, A.; Sato, N.; Shiomi, K.; Takahashi, Y.; et al. Anti-trypanosomal compound, sagamilactam, a new polyene macrocyclic lactam from Actinomadura sp. K13-0306. J. Antibiot. 2016, 69, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, Y.; Matsuoka, N.; In, Y.; Kataura, T.; Tashiro, E.; Saiki, I.; Sudoh, Y.; Duangmal, K.; Thamchaipenet, A. Nonthmicin, a Polyether Polyketide Bearing a Halogen-Modified Tetronate with Neuroprotective and Antiinvasive Activity from Actinomadura sp. Org. Lett. 2017, 19, 1406–1409. [Google Scholar] [CrossRef]
- Lu, C.; Xie, F.; Shan, C.; Shen, Y. Two novel cyclic hexapeptides from the genetically engineered Actinosynnema pretiosum. Appl. Microbiol. Biotechnol. 2017, 101, 2273–2279. [Google Scholar] [CrossRef]
- Frattaruolo, L.; Lacret, R.; Cappello, A.R.; Truman, A.W. A Genomics-Based Approach Identifies a Thioviridamide-Like Compound with Selective Anticancer Activity. ACS Chem. Biol. 2017, 12, 2815–2822. [Google Scholar] [CrossRef]
- Hoshino, S.; Ozeki, M.; Awakawa, T.; Morita, H.; Onaka, H.; Abe, I. Catenulobactins A and B, Heterocyclic Peptides from Culturing Catenuloplanes sp. with a Mycolic Acid-Containing Bacterium. J. Nat. Prod. 2018, 81, 2106–2110. [Google Scholar] [CrossRef]
- Pournejati, R.; Gust, R.; Kircher, B.; Karbalaei-Heidari, H.R. Microindoline 581, an Indole Derivative from Microbacterium Sp. RP581 as A Novel Selective Antineoplastic Agent to Combat Hepatic Cancer Cells: Production, Optimization and Structural Elucidation. Iran. J. Pharm. Res. 2020, 19, 290–305. [Google Scholar] [CrossRef]
- Fu, G.; Wang, R.; Ding, J.; Qi, H.; Zhao, Z.; Chen, C.; Zhang, H.; Xue, Z.; Wang, J.; Wu, M. Micromonospora zhangzhouensis sp. nov., a Novel Actinobacterium Isolated from Mangrove Soil, Exerts a Cytotoxic Activity in vitro. Sci. Rep. 2020, 10, 3889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmiento-Vizcaíno, A.; Braña, A.F.; Pérez-Victoria, I.; Martín, J.; De Pedro, N.; De La Cruz, M.; Díaz, C.; Vicente, F.; Acuña, J.L.; Reyes, F.; et al. Paulomycin G, a New Natural Product with Cytotoxic Activity against Tumor Cell Lines Produced by Deep-Sea Sediment Derived Micromonospora matsumotoense M-412 from the Avilés Canyon in the Cantabrian Sea. Mar. Drugs 2017, 15, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.J.; Zhang, S.Y.; Ye, Y.H.; Yu, Z.; Qi, H.; Zhang, H.; Xue, Z.L.; Wang, J.D.; Wu, M. Three New Isoflavonoid Glycosides from the Mangrove-Derived Actinomycete Micromonospora aurantiaca 110B. Mar. Drugs 2019, 17, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhao, W.; Jiang, H.L.; Zhou, J.; Chen, X.M.; Lian, Y.Y.; Jiang, H.; Lin, F. Rakicidins G-I, cyclic depsipeptides from marine Micromonospora chalcea FIM 02-523. Tetrahedron 2018, 74, 4151–4154. [Google Scholar] [CrossRef]
- Nie, Y.L.; Wu, Y.D.; Wang, C.X.; Lin, R.; Xie, Y.; Fang, D.S.; Jiang, H.; Lian, Y.Y. Structure elucidation and antitumour activity of a new macrolactam produced by marine-derived actinomycete Micromonospora sp. FIM05328. Nat. Prod. Res. 2018, 32, 2133–2138. [Google Scholar] [CrossRef]
- Gao, M.Y.; Qi, H.; Li, J.S.; Zhang, H.; Zhang, J.; Wang, J.D.; Xiang, W.S. A new naphthalenepropanoic acid analog from the marine-derived actinomycetes Micromonospora sp. HS-HM-036. J. Asian Nat. Prod. Res. 2017, 19, 930–934. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Chen, J.J.; Adhikari, A.; Yang, D.; Crnovcic, I.; Wang, N.; Chang, C.Y.; Rader, C.; Shen, B. Genome Mining of Micromonospora yangpuensis DSM 45577 as a Producer of an Anthraquinone-Fused Enediyne. Org. Lett. 2017, 19, 6192–6195. [Google Scholar] [CrossRef]
- Fukuda, T.; Takahashi, M.; Nagai, K.; Harunari, E.; Imada, C.; Tomoda, H. Isomethoxyneihumicin, a new cytotoxic agent produced by marine Nocardiopsis alba KM6-1. J. Antibiot. 2017, 70, 590–594. [Google Scholar] [CrossRef] [Green Version]
- Messaoudi, O.; Sudarman, E.; Bendahou, M.; Jansen, R.; Stadler, M.; Wink, J. Kenalactams A-E, Polyene Macrolactams Isolated from Nocardiopsis CG3. J. Nat. Prod. 2019, 82, 1081–1088. [Google Scholar] [CrossRef]
- Shaaban, K.A.; Shaaban, M.; Rahman, H.; Grün-Wollny, I.; Kämpfer, P.; Kelter, G.; Fiebig, H.H.; Laatsch, H. Karamomycins A-C: 2-Naphthalen-2-yl-thiazoles from Nonomuraea endophytica. J. Nat. Prod. 2019, 82, 870–877. [Google Scholar] [CrossRef]
- Yang, T.; Yamada, K.; Zhou, T.; Harunari, E.; Igarashi, Y.; Terahara, T.; Kobayashi, T.; Imada, C. Akazamicin, a cytotoxic aromatic polyketide from marine-derived Nonomuraea sp. J. Antibiot. 2019, 72, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Gamaleldin, N.M.; Bakeer, W.; Sayed, A.M.; Shamikh, Y.I.; El-Gendy, A.O.; Hassan, H.M.; Horn, H.; Abdelmohsen, U.R.; Hozzein, W.N. Exploration of Chemical Diversity and Antitrypanosomal Activity of Some Red Sea-Derived Actinomycetes Using the OSMAC Approach Supported by LC-MS-Based Metabolomics and Molecular Modelling. Antibiotics 2020, 9, 629. [Google Scholar] [CrossRef] [PubMed]
- Anh, C.V.; Kwon, J.-H.; Kang, J.S.; Lee, H.-S.; Heo, C.-S.; Shin, H.J. New Angucycline Glycosides from a Marine-Derived Bacterium Streptomyces ardesiacus. Int. J. Mol. Sci. 2022, 23, 13779. [Google Scholar] [CrossRef] [PubMed]
- Kaari, M.; Joseph, J.; Manikkam, R.; Kalyanasundaram, R.; Sivaraj, A.; Anbalmani, S.; Murthy, S.; Sahu, A.K.; Said, M.; Dastager, S.G. A Novel Finding: 2, 4-Di-tert-butylphenol from Streptomyces bacillaris ANS2 Effective Against Mycobacterium tuberculosis and Cancer Cell Lines. Appl. Biochem. Biotechnol. 2023, 195, 1–14. [Google Scholar] [CrossRef]
- Gui, C.; Yuan, J.; Mo, X.; Huang, H.; Zhang, S.; Gu, Y.C.; Ju, J. Cytotoxic Anthracycline Metabolites from a Recombinant Streptomyces. J. Nat. Prod. 2018, 81, 1278–1289. [Google Scholar] [CrossRef]
- Kifer, D.; Mužinić, V.; Klaric, M.Š. Antimicrobial potency of single and combined mupirocin and monoterpenes, thymol, menthol and 1,8-cineole against Staphylococcus aureus planktonic and biofilm growth. J. Antibiot. 2016, 69, 689–696. [Google Scholar] [CrossRef]
- Xiao, F.; Li, H.; Xu, M.; Li, T.; Wang, J.; Sun, C.; Hong, K.; Li, W. Staurosporine Derivatives Generated by Pathway Engineering in a Heterologous Host and Their Cytotoxic Selectivity. J. Nat. Prod. 2018, 81, 1745–1751. [Google Scholar] [CrossRef]
- Kaweewan, I.; Komaki, H.; Hemmi, H.; Hoshino, K.; Hosaka, T.; Isokawa, G.; Oyoshi, T.; Kodani, S. Isolation and structure determination of a new cytotoxic peptide, curacozole, from Streptomyces curacoi based on genome mining. J. Antibiot. 2019, 72, 1–7. [Google Scholar] [CrossRef]
- Ortiz-López, F.J.; Alcalde, E.; Sarmiento-Vizcaíno, A.; Díaz, C.; Cautain, B.; García, L.A.; Blanco, G.; Reyes, F. New 3-Hydroxyquinaldic Acid Derivatives from Cultures of the Marine Derived Actinomycete Streptomyces cyaneofuscatus M-157. Mar. Drugs 2018, 16, 371. [Google Scholar] [CrossRef] [Green Version]
- Kimata, S.; Matsuda, T.; Suizu, Y.; Hayakawa, Y. Prodigiosin R2, a new prodigiosin from the roseophilin producer Streptomyces griseoviridis 2464-S5. J. Antibiot. 2018, 71, 393–396. [Google Scholar] [CrossRef]
- Liu, S.H.; Wang, W.; Wang, K.B.; Zhang, B.; Li, W.; Shi, J.; Jiao, R.H.; Tan, R.X.; Ge, H.M. Heterologous Expression of a Cryptic Giant Type I PKS Gene Cluster Leads to the Production of Ansaseomycin. Org. Lett. 2019, 21, 3785–3788. [Google Scholar] [CrossRef] [PubMed]
- Koomsiri, W.; Inahashi, Y.; Kimura, T.; Shiomi, K.; Takahashi, Y.; Omura, S.; Thamchaipenet, A.; Nakashima, T. Bisoxazolomycin A: A new natural product from ‘Streptomyces subflavus subsp. irumaensis’ AM-3603. J. Antibiot. 2017, 70, 1142–1145. [Google Scholar] [CrossRef]
- Zheng, D.; Ding, N.; Jiang, Y.; Zhang, J.; Ma, J.; Chen, X.; Liu, J.; Han, L.; Huang, X. Albaflavenoid, a new tricyclic sesquiterpenoid from Streptomyces violascens. J. Antibiot. 2016, 69, 773–775. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Han, L.; Qu, X.; Chen, X.; Zhong, J.; Bi, X.; Liu, J.; Jiang, Y.; Jiang, C.; Huang, X. Cytotoxic Fusicoccane-Type Diterpenoids from Streptomyces violascens Isolated from Ailuropoda melanoleuca Feces. J. Nat. Prod. 2017, 80, 837–844. [Google Scholar] [CrossRef]
- Lv, Q.; Fan, Y.; Tao, G.; Fu, P.; Zhai, J.; Ye, B.; Zhu, W. Sekgranaticin, a SEK34b-Granaticin Hybrid Polyketide from Streptomyces sp. 166. J. Org Chem. 2019, 84, 9087–9092. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, L.; Ye, X.; Anjum, K.; Lian, X.Y.; Zhang, Z. New streptophenazines from marine Streptomyces sp. 182SMLY. Nat. Prod. Res. 2017, 31, 411–417. [Google Scholar] [CrossRef]
- Liu, C.X.; Liu, S.H.; Zhao, J.W.; Zhang, J.; Wang, X.J.; Li, J.S.; Wang, J.D.; Xiang, W.S. A new spectinabilin derivative with cytotoxic activity from ant-derived Streptomyces sp. 1H-GS5. J. Asian Nat. Prod. Res. 2017, 19, 924–929. [Google Scholar] [CrossRef]
- Zhou, B.; Qin, L.L.; Ding, W.J.; Ma, Z.J. Cytotoxic indolocarbazoles alkaloids from the Streptomyces sp. A65. Tetrahedron 2018, 74, 726–730. [Google Scholar] [CrossRef]
- Wang, X.; Elshahawi, S.I.; Ponomareva, L.V.; Ye, Q.; Liu, Y.; Copley, G.C.; Hower, J.C.; Hatcher, B.E.; Kharel, M.K.; Van Lanen, S.G.; et al. Structure Determination, Functional Characterization, and Biosynthetic Implications of Nybomycin Metabolites from a Mining Reclamation Site-Associated Streptomyces. J. Nat. Prod. 2019, 82, 3469–3476. [Google Scholar] [CrossRef]
- Saito, S.; Fujimaki, T.; Panbangred, W.; Sawa, R.; Igarashi, Y.; Imoto, M. Antarlides F–H, new members of the antarlide family produced by Streptomyces sp. BB47. J. Antibiot. 2017, 70, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Liu, Z.; Zhang, Z.; Zhang, X.; Zhu, T.; Gu, Q.; Li, W.; Che, Q.; Li, D. Geranylpyrrol A and Piericidin F from Streptomyces sp. CHQ-64 ΔrdmF. J. Nat. Prod. 2017, 80, 1684–1687. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.N.; Zhang, H.J.; Li, J.Q.; Ding, W.J.; Ma, Z.J. Bioactive Indolocarbazoles from the Marine-Derived Streptomyces sp. DT-A61. J. Nat. Prod. 2018, 81, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, M.S.; Elmallah, M.I.Y.; Mohamed, A.A.; Ishibashi, M. Sharkquinone, a new ana-quinonoid tetracene derivative from marine-derived Streptomyces sp. EGY1 with TRAIL resistance-overcoming activity. J. Nat. Med. 2017, 71, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Lu, M.C.; Chung, H.M.; Weng, C.F.; Su, J.H.; Yang, Y.T.; Su, Y.D.; Chang, Y.C.; Kuo, J.; Wu, Y.C.; et al. Bafilomycin M, a new cytotoxic bafilomycin produced by a Streptomyces sp. isolated from a marine sponge Theonella sp. Tetrahedron Lett. 2016, 57, 4863–4865. [Google Scholar] [CrossRef]
- Chen, H.; Cai, K.; Yao, R. A new macrolactam derivative from the marine actinomycete HF-11225. J. Antibiot. 2018, 71, 477–479. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.J.; Li, J.Q.; Zhang, H.J.; Ding, W.J.; Ma, Z.J. Cyclizidine-Type Alkaloids from Streptomyces sp. HNA39. J. Nat. Prod. 2018, 81, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Li, J.M.; Qi, H.; Zhang, H.; Zhang, J.; Xiang, W.S.; Wang, J.D.; Wang, X.J. Two new lankacidin-related metabolites from Streptomyces sp. HS-NF-1178. J. Antibiot. 2018, 71, 397–401. [Google Scholar] [CrossRef]
- Zhao, X.L.; Wang, H.; Xue, Z.L.; Li, J.S.; Qi, H.; Zhang, H.; Zhao, T.; Wang, J.D.; Xiang, W.S. Two new glutarimide antibiotics from Streptomyces sp. HS-NF-780. J. Antibiot. 2019, 72, 241–245. [Google Scholar] [CrossRef]
- Yixizhuoma; Ishikawa, N.; Abdelfattah, M.S.; Ishibashi, M. Elmenols C-H, new angucycline derivatives isolated from a culture of Streptomyces sp. IFM 11490. J. Antibiot. 2017, 70, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Son, S.; Jang, M.; Lee, B.; Hong, Y.S.; Ko, S.K.; Jang, J.H.; Ahn, J.S. Ulleungdin, a Lasso Peptide with Cancer Cell Migration Inhibitory Activity Discovered by the Genome Mining Approach. J. Nat. Prod. 2018, 81, 2205–2211. [Google Scholar] [CrossRef]
- Son, S.; Ko, S.K.; Jang, M.; Lee, J.K.; Kwon, M.C.; Kang, D.H.; Ryoo, I.J.; Lee, J.S.; Hong, Y.S.; Kim, B.Y.; et al. Polyketides and Anthranilic Acid Possessing 6-Deoxy-α-l-talopyranose from a Streptomyces Species. J. Nat. Prod. 2017, 80, 1378–1386. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.X.; Hou, G.X.; Luo, J.; Yang, J.; Yan, Y.; Huang, S.X. New phenoxazinone-related alkaloids from strain Streptomyces sp. KIB-H1318. J. Antibiot. 2018, 71, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Cao, P.; Ma, Y.T.; Luo, J.; Yan, Y.; Li, R.T.; Huang, S.X. A new actinomycin Z analogue with an additional oxygen bridge between chromophore and β-depsipentapeptide from Streptomyces sp. KIB-H714. Nat. Prod. Res. 2019, 33, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Xie, Z.P.; Yang, Q.; Feng, L.L.; Zhang, L.; Zhang, Y.Z.; Li, X.N.; Pescitelli, G.; Zhang, S.M. Kiamycins B and C, unusual bridged angucyclinones from a marine sediment-derived Streptomyces sp. Tetrahedron Lett. 2018, 59, 2176–2180. [Google Scholar] [CrossRef]
- Kawahara, T.; Izumikawa, M.; Kozone, I.; Hashimoto, J.; Kagaya, N.; Koiwai, H.; Komatsu, M.; Fujie, M.; Sato, N.; Ikeda, H.; et al. Neothioviridamide, a Polythioamide Compound Produced by Heterologous Expression of a Streptomyces sp. Cryptic RiPP Biosynthetic Gene Cluster. J. Nat. Prod. 2018, 81, 264–269. [Google Scholar] [CrossRef]
- Lu, D.D.; Ren, J.W.; Du, Q.Q.; Song, Y.J.; Lin, S.Q.; Li, X.; Li, E.W.; Xie, W.D. p-Terphenyls and actinomycins from a Streptomyces sp. associated with the larva of mud dauber wasp. Nat. Prod. Res. 2021, 35, 1869–1873. [Google Scholar] [CrossRef]
- Song, Y.J.; Zheng, H.B.; Peng, A.H.; Ma, J.H.; Lu, D.D.; Li, X.; Zhang, H.Y.; Xie, W.D. Strepantibins A-C: Hexokinase II Inhibitors from a Mud Dauber Wasp Associated Streptomyces sp. J. Nat. Prod. 2019, 82, 1114–1119. [Google Scholar] [CrossRef]
- Cheng, P.; Xu, K.; Chen, Y.C.; Wang, T.T.; Chen, Y.; Yang, C.L.; Ma, S.Y.; Liang, Y.; Ge, H.M.; Jiao, R.H. Cytotoxic aromatic polyketides from an insect derived Streptomyces sp. NA4286. Tetrahedron Lett. 2019, 60, 1706–1709. [Google Scholar] [CrossRef]
- Lu, C.; Zhao, Y.; Jia, W.-Q.; Zhang, H.; Qi, H.; Xiang, W.-S.; Wang, J.-D.; Wang, X.-J. A new anthracycline-type metabolite from Streptomyces sp. NEAU-L3. J. Antibiot. 2017, 70, 1026–1028. [Google Scholar] [CrossRef]
- Kawahara, T.; Fujiwara, T.; Kagaya, N.; Shin-Ya, K. JBIR-150, a novel 20-membered polyene macrolactam from marine-derived Streptomyces sp. OPMA00071. J. Antibiot. 2018, 71, 390–392. [Google Scholar] [CrossRef]
- Abbas, M.; Elshahawi, S.I.; Wang, X.; Ponomareva, L.V.; Sajid, I.; Shaaban, K.A.; Thorson, J.S. Puromycins B-E, Naturally Occurring Amino-Nucleosides Produced by the Himalayan Isolate Streptomyces sp. PU-14G. J. Nat. Prod. 2018, 81, 2560–2566. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chai, W.; Wang, W.; Song, T.; Lian, X.Y.; Zhang, Z. Cytotoxic Bagremycins from Mangrove-Derived Streptomyces sp. Q22. J. Nat. Prod. 2017, 80, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, Y.; Akimoto, M.; Ishikawa, A.; Izawa, M.; Shin-Ya, K. Curromycin A as a GRP78 downregulator and a new cyclic dipeptide from Streptomyces sp. J. Antibiot. 2016, 69, 187–188. [Google Scholar] [CrossRef] [PubMed]
- Nogawa, T.; Okano, A.; Lim, C.L.; Futamura, Y.; Shimizu, T.; Takahashi, S.; Ibrahim, D.; Osada, H. Opantimycin A, a new metabolite isolated from Streptomyces sp. RK88-1355. J. Antibiot. 2017, 70, 222–225. [Google Scholar] [CrossRef]
- Cheng, C.; Othman, E.M.; Fekete, A.; Krischke, M.; Stopper, H.; Edrada-Ebel, R.A.; Mueller, M.J.; Hentschel, U.; Abdelmohsen, U.R. Strepoxazine A, a new cytotoxic phenoxazin from the marine sponge-derived bacterium Streptomyces sp. SBT345. Tetrahedron Lett. 2016, 57, 4196–4199. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ma, L.; Zhang, L.; Zhang, W.; Zhu, Y.; Chen, Y.; Zhang, W.; Zhang, C. Functional characterization of the halogenase SpmH and discovery of new deschloro-tryptophan dimers. Org. Biomol. Chem. 2019, 17, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Zhang, Q.; Jiang, X.; Ma, L.; Long, T.; Cheng, Z.; Zhang, C.; Zhu, Y. New piericidin derivatives from the marine-derived Streptomyces sp. SCSIO 40063 with cytotoxic activity. Nat. Prod. Res. 2022, 36, 2458–2464. [Google Scholar] [CrossRef]
- Jiang, Y.J.; Gan, L.S.; Ding, W.J.; Chen, Z.; Ma, Z.J. Cytotoxic gephyromycins from the Streptomyces sp. SS13I. Tetrahedron Lett. 2017, 58, 3747–3750. [Google Scholar] [CrossRef]
- Zhang, D.; Jiang, Y.; Li, J.; Ding, W.; Chen, Z.; Ma, Z. Thioquinomycins A-D, novel naphthothiophenediones from the marine-derived Streptomyces sp. SS17F. Tetrahedron 2018, 74, 6150–6154. [Google Scholar] [CrossRef]
- Xu, C.D.; Zhang, H.J.; Ma, Z.J. Pyrimidine Nucleosides from Streptomyces sp. SSA28. J. Nat. Prod. 2019, 82, 2509–2516. [Google Scholar] [CrossRef]
- Lee, B.; Lee, G.-E.; Hwang, G.J.; Heo, K.T.; Lee, J.K.; Jang, J.-P.; Hwang, B.Y.; Jang, J.-H.; Cho, Y.-Y.; Hong, Y.-S. Rubiflavin G, photorubiflavin G, and photorubiflavin E: Novel pluramycin derivatives from Streptomyces sp. W2061 and their anticancer activity against breast cancer cells. J. Antibiot. 2023, 76, 1–7. [Google Scholar] [CrossRef]
- Xu, Y.; Hu, B.Y.; Qin, Y.; Zhuang, L.; Yang, Y.B.; Zhao, L.X. Jiangchuanmycin, a New Pyrrolizidine Analog from Streptomyces sp. YIM S01863. Chem. Biodivers. 2023, 20, e202201240. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, Y.; Yang, Y.; Chen, H. Shellmycin A-D, Novel Bioactive Tetrahydroanthra-γ-Pyrone Antibiotics from Marine Streptomyces sp. Shell-016. Mar. Drugs 2020, 18, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Sayed, K.A. Natural Products as Antiviral Agents. Stud. Nat. Prod. Chem. 2000, 24, 473–572. [Google Scholar] [CrossRef]
- Patridge, E.; Gareiss, P.; Kinch, M.S.; Hoyer, D. An analysis of FDA-approved drugs: Natural products and their derivatives. Drug Discov. Today 2016, 21, 204–207. [Google Scholar] [CrossRef]
- Farmer, P.B.; Suhadolnik, R.J. Nucleoside antibiotics. Biosynthesis of arabinofuranosyladenine by Streptomyces antibioticus. Biochemistry 1972, 11, 911–916. [Google Scholar] [CrossRef]
- Sayed, A.M.; Alhadrami, H.A.; El-Gendy, A.O.; Shamikh, Y.I.; Belbahri, L.; Hassan, H.M.; Abdelmohsen, U.R.; Rateb, M.E. Microbial Natural Products as Potential Inhibitors of SARS-CoV-2 Main Protease (Mpro). Microorganisms 2020, 8, 970. [Google Scholar] [CrossRef]
- Raveh, A.; Delekta, P.C.; Dobry, C.J.; Peng, W.; Schultz, P.J.; Blakely, P.K.; Tai, A.W.; Matainaho, T.; Irani, D.N.; Sherman, D.H.; et al. Discovery of potent broad spectrum antivirals derived from marine actinobacteria. PLoS ONE 2013, 8, e82318. [Google Scholar] [CrossRef] [Green Version]
- Jakubiec-Krzesniak, K.; Rajnisz-Mateusiak, A.; Guspiel, A.; Ziemska, J.; Solecka, J. Secondary Metabolites of Actinomycetes and their Antibacterial, Antifungal and Antiviral Properties. Pol. J. Microbiol. 2018, 67, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Rawal, K.R.; Bariwal, J.; Singh, V. Chemistry and Bioactivities of Aristeromycins: An Overview. Curr. Top. Med. Chem. 2016, 16, 3258–3273. [Google Scholar] [CrossRef]
- Selim, M.S.M.; Abdelhamid, S.A.; Mohamed, S.S. Secondary metabolites and biodiversity of actinomycetes. J. Genet. Eng. Biotechnol. 2021, 19, 72. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Bayer, K.; Hentschel, U. Diversity, abundance and natural products of marine sponge-associated actinomycetes. Nat. Prod. Rep. 2014, 31, 381–399. [Google Scholar] [CrossRef] [PubMed]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antiviral Res. 2020, 178, 104787. [Google Scholar] [CrossRef] [PubMed]
- Euanorasetr, J.; Intra, B.; Thunmrongsiri, N.; Limthongkul, J.; Ubol, S.; Anuegoonpipat, A.; Kurosu, T.; Ikuta, K.; Nihira, T.; Panbangred, W. In vitro antiviral activity of spirotetronate compounds against dengue virus serotype 2. J. Gen. Appl. Microbiol. 2019, 65, 197–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, L.; Oueis, E.; Kaur, A.; Safaei, N.; Kirsch, S.H.; Gunesch, A.P.; Haid, S.; Rand, U.; Čičin-Šain, L.; Fu, C. Persicamidines—Unprecedented Sesquarterpenoids with Potent Antiviral Bioactivity against Coronaviruses. Angew. Chem. Int. Ed. 2023, 62, e202214595. [Google Scholar] [CrossRef]
- Shuai, H.; Myronovskyi, M.; Nadmid, S.; Luzhetskyy, A. Identification of a Biosynthetic Gene Cluster Responsible for the Production of a New Pyrrolopyrimidine Natural Product-Huimycin. Biomolecules 2020, 10, 1074. [Google Scholar] [CrossRef]
- Zhang, Y.; Fang, W.; Wang, K.; Zhang, Z.; Wu, Z.; Shi, L.; Liu, F.; Wan, Z.; Liu, M. Napyradiomycin A4 and Its Relate Compounds, a New Anti-PRV Agent and Their Antibacterial Activities, from Streptomyces kebangsaanensis WS-68302. Molecules 2023, 28, 640. [Google Scholar] [CrossRef]
- Liu, M.; Ren, M.; Zhang, Y.; Wan, Z.; Wang, Y.; Wu, Z.; Wang, K.; Fang, W.; Yang, X. Antiviral activity of benzoheterocyclic compounds from soil-derived Streptomyces jiujiangensis NBERC-24992. Molecules 2023, 28, 878. [Google Scholar] [CrossRef]
- Hao, X.; Li, S.; Wang, G.; Li, J.; Peng, Z.; Zhang, Y.; Yu, L.; Gan, M. Zelkovamycins F and G, Cyclopeptides with Cα-Methyl-threonine Residues, from an Endophytic Kitasatospora sp. J. Nat. Prod. 2022, 85, 1715–1722. [Google Scholar] [CrossRef]
- Kimura, T.; Suga, T.; Kameoka, M.; Ueno, M.; Inahashi, Y.; Matsuo, H.; Iwatsuki, M.; Shigemura, K.; Shiomi, K.; Takahashi, Y.; et al. New tetrahydroquinoline and indoline compounds containing a hydroxy cyclopentenone, virantmycin B and C, produced by Streptomyces sp. AM-2504. J. Antibiot. 2019, 72, 169–173. [Google Scholar] [CrossRef]
- Kim, S.H.; Ha, T.K.Q.; Oh, W.K.; Shin, J.; Oh, D.C. Antiviral Indolosesquiterpenoid Xiamycins C-E from a Halophilic Actinomycete. J. Nat. Prod. 2016, 79, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wang, J.; Tang, Y.; Guo, Z.; Bai, J.; Wu, L.; Su, J.; Cen, S.; Yu, L.; Zhang, D. Geninthiocins E and F, two new cyclic thiopeptides with antiviral activities from soil-derived Streptomyces sp. CPCC 200267 using OSMAC strategy. J. Antibiot. 2023, 76, 101–104. [Google Scholar] [CrossRef]
- Li, F.; Lu, S.; Xie, X.; Fan, S.; Chen, D.; Wu, S.; He, J. Antiviral properties of extracts of Streptomyces sp. SMU 03 isolated from the feces of Elephas maximus. Fitoterapia 2020, 143, 104600. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Funayama, K.; Kato, W.; Okuda, M.; Kawamoto, M.; Matsubara, T.; Sato, T.; Sato, A.; Otsuguro, S.; Sasaki, M. Dihydromaniwamycin E, a Heat-Shock Metabolite from Thermotolerant Streptomyces sp. JA74, Exhibiting Antiviral Activity against Influenza and SARS-CoV-2 Viruses. J. Nat. Prod. 2022, 85, 2583–2591. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, Y.; Yu, L.; Ikeda, M.; Oikawa, T.; Kitani, S.; Nihira, T.; Bayanmunkh, B.; Panbangred, W. Jomthonic Acid A, a Modified Amino Acid from a Soil-Derived Streptomyces. J. Nat. Prod. 2012, 75, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Oikawa, T.; Kitani, S.; Nihira, T.; Bayanmunkh, B.; Panbangred, W.; Igarashi, Y. Jomthonic acids B and C, two new modified amino acids from Streptomyces sp. J. Antibiot. 2014, 67, 345–347. [Google Scholar] [CrossRef] [Green Version]
- Inahashi, Y.; Iwatsuki, M.; Ishiyama, A.; Matsumoto, A.; Hirose, T.; Oshita, J.; Sunazuka, T.; Panbangred, W.; Takahashi, Y.; Kaiser, M.; et al. Actinoallolides A–E, New Anti-trypanosomal Macrolides, Produced by an Endophytic Actinomycete, Actinoallomurus fulvus MK10-036. Org. Lett. 2015, 17, 864–867. [Google Scholar] [CrossRef]
- Akiyama, H.; Oku, N.; Harunari, E.; Panbangred, W.; Igarashi, Y. Complete NMR assignment and absolute configuration of k4610422, a norditerpenoid inhibitor of testosterone-5α-reductase originally from Streptosporangium: Rediscovery from a thermophilic Actinomadura. J. Antibiot. 2020, 73, 60–65. [Google Scholar] [CrossRef]
- Lyddiard, D.; Jones, G.L.; Greatrex, B.W. Keeping it simple: Lessons from the golden era of antibiotic discovery. FEMS Microbiol. Lett. 2016, 363, 84. [Google Scholar] [CrossRef] [Green Version]
- Majumder, M.A.A.; Rahman, S.; Cohall, D.; Bharatha, A.; Singh, K.; Haque, M.; Gittens-St Hilaire, M. Antimicrobial Stewardship: Fighting Antimicrobial Resistance and Protecting Global Public Health. Infect. Drug Resist. 2020, 13, 4713. [Google Scholar] [CrossRef]
- Baltz, R.H. Renaissance in antibacterial discovery from actinomycetes. Curr. Opin. Pharm. 2008, 8, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Lazzarini, A.; Cavaletti, L.; Toppo, G.; Marinelli, F. Rare genera of actinomycetes as potential producers of new antibiotics. Antonie Van Leeuwenhoek 2000, 78, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Kohli, I.; Joshi, N.C.; Mohapatra, S.; Varma, A. Extremophile—An Adaptive Strategy for Extreme Conditions and Applications. Curr. Genom. 2020, 21, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Subramani, R.; Aalbersberg, W. Culturable rare Actinomycetes: Diversity, isolation and marine natural product discovery. Appl. Microbiol. Biotechnol. 2013, 97, 9291–9321. [Google Scholar] [CrossRef]
- Mahajan, G.B.; Balachandran, L. Sources of antibiotics: Hot springs. Biochem. Pharmacol. 2017, 134, 35–41. [Google Scholar] [CrossRef]
- Thawai, C.; Thamsathit, W.; Kudo, T. Planosporangium thailandense sp. nov., isolated from soil from a Thai hot spring. Int. J. Syst. Evol. Microbiol. 2013, 63, 1051–1055. [Google Scholar] [CrossRef] [Green Version]
- Bull, A.T.; Idris, H.; Sanderson, R.; Asenjo, J.; Andrews, B.; Goodfellow, M. High altitude, hyper-arid soils of the Central-Andes harbor mega-diverse communities of actinobacteria. Extremophiles 2017, 22, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Shi, Y.-L.; Wang, H.; Zhang, T.; Yu, L.-Y.; Sun, H.; Zhang, Y.-Q. Diversity of Bacteria and the Characteristics of Actinobacteria Community Structure in Badain Jaran Desert and Tengger Desert of China. Front. Microbiol. 2018, 9, 1068. [Google Scholar] [CrossRef] [Green Version]
- Li, W.J.; Zhang, Y.G.; Zhang, Y.Q.; Tang, S.K.; Xu, P.; Xu, L.H.; Jiang, C.L. Streptomyces sodiiphilus sp. nov., a novel alkaliphilic actinomycete. Int. J. Syst. Evol. Microbiol. 2005, 55, 1329–1333. [Google Scholar] [CrossRef] [Green Version]
- Hui, M.L.-Y.; Tan, L.T.H.; Letchumanan, V.; He, Y.-W.; Fang, C.-M.; Chan, K.-G.; Law, J.W.F.; Lee, L.H. The Extremophilic Actinobacteria: From Microbes to Medicine. Antibiotics 2021, 10, 682. [Google Scholar] [CrossRef]
- Singh, R.; Dubey, A.K. Diversity and Applications of Endophytic Actinobacteria of Plants in Special and Other Ecological Niches. Front. Microbiol. 2018, 9, 1767. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Chater, K.F.; Cerdeno-Tarraga, A.M.; Chalis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Hoskisson, P.A.; van Wezel, G.P. Streptomyces coelicolor. Trends Microbiol. 2019, 27, 468–469. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Weber, T.; Rausch, C.; Lopez, P.; Hoof, I.; Gaykova, V.; Huson, D.H.; Wohlleben, W. CLUSEAN: A computer-based framework for the automated analysis of bacterial secondary metabolite biosynthetic gene clusters. J. Biotechnol. 2009, 140, 13–17. [Google Scholar] [CrossRef]
- Skinnider, M.A.; Johnston, C.W.; Gunabalasingam, M.; Merwin, N.J.; Kieliszek, A.M.; MacLellan, R.J.; Li, H.; Ranieri, M.R.M.; Webster, A.L.H.; Cao, M.P.T.; et al. Comprehensive prediction of secondary metabolite structure and biological activity from microbial genome sequences. Nat. Comm. 2020, 11, 6058. [Google Scholar] [CrossRef]
- Li, M.H.T.; Ung, P.M.U.; Zajkowski, J.; Garneau-Tsodikova, S.; Sherman, D.H. Automated genome mining for natural products. BMC Bioinform. 2009, 10, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starcevic, A.; Zucko, J.; Simunkovic, J.; Long, P.F.; Cullum, J.; Hranueli, D. ClustScan: An integrated program package for the semi-automatic annotation of modular biosynthetic gene clusters and in silico prediction of novel chemical structures. Nucleic Acids Res. 2008, 36, 6882–6892. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Jiang, W.; Lu, Y. New strategies and approaches for engineering biosynthetic gene clusters of microbial natural products. Biotechnol. Adv. 2017, 35, 936–949. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, Y.; Huang, C.; Luo, Y. Recent Advances in Silent Gene Cluster Activation in Streptomyces. Front. Bioeng. Biotechnol. 2021, 9, 88. [Google Scholar] [CrossRef]
- Chou, W.K.W.; Fanizza, I.; Uchiyama, T.; Komatsu, M.; Ikeda, H.; Cane, D.E. Genome mining in Streptomyces avermitilis: Cloning and characterization of SAV_76, the synthase for a new sesquiterpene, avermitilol. J. Am. Chem. Soc. 2010, 132, 8850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring structural diversity of microbe secondary metabolites using OSMAC strategy: A literature review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef] [Green Version]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asamizu, S.; Ozaki, T.; Teramoto, K.; Satoh, K.; Onaka, H. Killing of Mycolic Acid-Containing Bacteria Aborted Induction of Antibiotic Production by Streptomyces in Combined-Culture. PLoS ONE 2015, 10, e0142372. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Li, X.; Li, Z.; Zhan, X.; Mao, X.; Li, Y. The Application of Regulatory Cascades in Streptomyces: Yield Enhancement and Metabolite Mining. Front. Microbiol. 2020, 11, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tawfike, A.; Attia, E.Z.; Desoukey, S.Y.; Hajjar, D.; Makki, A.A.; Schupp, P.J.; Edrada-Ebel, R.A.; Abdelmohsen, U.R. New bioactive metabolites from the elicited marine sponge-derived bacterium Actinokineospora spheciospongiae sp. nov. AMB Express 2019, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Li, Y.; Banakar, S.P.; Liu, L.; Shao, C.; Li, Z.; Wang, C. New metabolites from the Co-culture of marine-derived actinomycete Streptomyces rochei MB037 and fungus Rhinocladiella similis 35. Front. Microbiol. 2019, 10, 915. [Google Scholar] [CrossRef] [Green Version]
- Tyurin, A.P.; Alferova, V.A.; Korshun, V.A. Chemical Elicitors of Antibiotic Biosynthesis in Actinomycetes. Microorganisms 2018, 6, 52. [Google Scholar] [CrossRef] [Green Version]
- Zong, G.; Fu, J.; Zhang, P.; Zhang, W.; Xu, Y.; Cao, G.; Zhang, R. Use of elicitors to enhance or activate the antibiotic production in Streptomyces. Crit. Rev. Biotechnol. 2022, 42, 1260–1283. [Google Scholar] [CrossRef]
- Choi, S.U.; Lee, C.K.; Hwang, Y.I.; Kinosita, H.; Nihira, T. γ-Butyrolactone autoregulators and receptor proteins in non-Streptomyces actinomycetes producing commercially important secondary metabolites. Arch. Microbiol. 2003, 180, 303–307. [Google Scholar] [CrossRef]
- Onaka, H. Novel antibiotic screening methods to awaken silent or cryptic secondary metabolic pathways in actinomycetes. J. Ant. 2017, 70, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Dashti, Y.; Grkovic, T.; Abdelmohsen, U.R.; Hentschel, U.; Quinn, R.J. Actinomycete Metabolome Induction/Suppression with N-Acetylglucosamine. J. Nat. Prod. 2017, 80, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Mast, Y.; Stegmann, E. Actinomycetes: The Antibiotics Producers. Antibiotics 2019, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Jose, P.A.; Jha, B. New Dimensions of Research on Actinomycetes: Quest for Next Generation Antibiotics. Front. Microbiol. 2016, 7, 1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadelhak, G.G.; El-Tarabily, K.A.; Al-Kaabi, F.K. Insect Control Using Chitinolytic Soil Actinomycetes as Bio control Agents. Int. J. Agri. Biol. 2005, 7, 627–633. [Google Scholar]
- Lam, K.S. Discovery of novel metabolites from marine actinomycetes. Curr. Opin. Microbiol. 2006, 9, 245–251. [Google Scholar] [CrossRef]
- Vijayabharathi, R.; Kumari, B.R.; Sathya, A.; Srinivas, V.; Abhishek, R.; Sharma, H.; Gopalakrishnan, S. Biological activity of entomopathogenic actinomycetes against lepidopteran insects (Noctuidae: Lepidoptera). Can. J. Plant Sci. 2014, 94, 759–769. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.-S.; Kim, H.-J.; Lee, H.-S.; Kim, P.; Kim, E.-S. Genome mining of rare actinomycetes and cryptic pathway awakening. Process. Biochem. 2015, 50, 1184–1193. [Google Scholar] [CrossRef]
- Peng, Q.; Gao, G.; Lü, J.; Long, Q.; Chen, X.; Zhang, F.; Xu, M.; Liu, K.; Wang, Y.; Deng, Z.; et al. Engineered Streptomyces lividans Strains for Optimal Identification and Expression of Cryptic Biosynthetic Gene Clusters. Front. Microbiol. 2018, 9, 3042. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.; Hwang, S.; Kim, J.; Cho, S.; Palsson, B.; Cho, B.K. Mini review: Genome mining approaches for the identification of secondary metabolite biosynthetic gene clusters in Streptomyces. Comput. Struct. Biotechnol. J. 2020, 18, 1548–1556. [Google Scholar] [CrossRef]
- Gomez-Escribano, J.P.; Bibb, M.J. Engineering Streptomyces coelicolor for heterologous expression of secondary metabolite gene clusters. Microb. Biotechnol. 2011, 4, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, D.; Okada, B.K.; Wu, Y.; Xu, F.; Seyedsayamdost, M.R. Recent advances in activating silent biosynthetic gene clusters in bacteria. Curr. Opin. Microbiol. 2018, 45, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, R.G.; Olano, C.; Gómez, C.; Fernández, R.; Braña, A.F.; Méndez, C.; Calle, F.; Salas, J.A. Characterization and engineering of the biosynthesis gene cluster for antitumor macrolides PM100117 and PM100118 from a marine actinobacteria: Generation of a novel improved derivative. Microb. Cell Factories 2016, 15, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, S.; Onaka, H.; Abe, I. Activation of silent biosynthetic pathways and discovery of novel secondary metabolites in actinomycetes by co-culture with mycolic acid-containing bacteria. J. Ind. Microbiol. Biotechnol. 2018, 46, 363–374. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The life cycle of actinomycetes, starting from conidiospores until sporulation. The various types of conidiospores are shown in the expanded box.
Figure 1.
The life cycle of actinomycetes, starting from conidiospores until sporulation. The various types of conidiospores are shown in the expanded box.

Figure 2.
Diversity of actinomycetes habitats. Actinomycetes are predominantly found in various ecological habitats e.g., marine ecosystems (of water bodies, coral reefs, seawater, mangrove forest), and terrestrial ecosystems (soil and plants, and insects).
Figure 2.
Diversity of actinomycetes habitats. Actinomycetes are predominantly found in various ecological habitats e.g., marine ecosystems (of water bodies, coral reefs, seawater, mangrove forest), and terrestrial ecosystems (soil and plants, and insects).

Figure 3.
Overview of genome mining approaches. BGCs and their metabolites are predicted from genome sequence data and confirmed by cloning the gene cluster either alone or in a combinational BGC into a heterologous host. Then, liquid chromatography with tandem mass spectrometry (LC-MS/MS) database can be used to predict the compound’s type, and its bioactivity should be confirmed.
Figure 3.
Overview of genome mining approaches. BGCs and their metabolites are predicted from genome sequence data and confirmed by cloning the gene cluster either alone or in a combinational BGC into a heterologous host. Then, liquid chromatography with tandem mass spectrometry (LC-MS/MS) database can be used to predict the compound’s type, and its bioactivity should be confirmed.

Figure 4.
The OSMAC methodology (Inspired by the overview of OSMAC in the review of Romano et al., 2018 [317]). (a) Chemical elicitors can trigger silent compounds, (b) changing nutrient regimes, (c) altering physical parameters such as temperature, and (d) Co-cultivation. These methods can improve and trigger the production of many secondary metabolites.
Figure 4.
The OSMAC methodology (Inspired by the overview of OSMAC in the review of Romano et al., 2018 [317]). (a) Chemical elicitors can trigger silent compounds, (b) changing nutrient regimes, (c) altering physical parameters such as temperature, and (d) Co-cultivation. These methods can improve and trigger the production of many secondary metabolites.

Table 1.
Examples of antibacterial metabolites identified from Actinomycetota from 2016 to 2023 *.
| Organism (s) | Compound Name (s) | Reference (s) |
|---|---|---|
| Actinoallomurus sp. ID145113, 145206, 145754 | Paramagnetoquinones A–C | [66] |
| Actinomadura atramentaria NBRC 14695 | Cinnamycin B | [67] |
| Actinomadura sp. KC191 | Actinomadurol | [68] |
| Amycolatopsis sp. IRD-009 | Pradimicin-IRD | [69] |
| Amycolatopsis sp. M39 | Macrotermycins A & C | [70] |
| Amycolatopsis sp. MCC0218 | Enceleamycins A–C | [71] |
| Amycolatopsis sp. ML1-hF4 | Pargamicins B–D | [72] |
| Kibdelosporangium phytohabitans XY-R10 | Maipomycin A (MaiA) | [73] |
| Kitasatospora sp. MG372-hF19 | Biospolides C-E | [74] |
| Kocuria marina CMG S2 | Kocumarin | [75] |
| Micromonospora carbonacea LS276 | Tetrocarcin Q | [76,77] |
| Micromonospora harpali SCSIO GJ089 | Icrosporanate A, Tetrocarcin P, Microsporanates B–F | [78] |
| Micromonospora chalcea FIM 02 | Rakicidins G–I | [79] |
| Micromonospora sp. UR56 + Actinokineospora sp. EG49 | Phenazine-1-carboxylic acid, Aestivophoenin C, Methyl saphenate | [80] |
| Micromonospora sp. WMMB-235 + Rhodococcus sp. WMMA-185 | Keyicin | [81] |
| Micromonospora sp. 5-297 | Tetrocarcins N | [82] |
| Micromonospora sp. CA-214671 | Phocoenamicin B | [83] |
| Micromonospora sp. RJA4480 | Sporalactam B | [84] |
| Micromonospora sp. SCSIO 07395 | Microechmycin A | [85] |
| Micromonospora sp. TP-A0468 | 16-demethylrifamycin S | [86] |
| Micromonospora endolithica | loseolamycins A1, A2 | [87] |
| Micromonospora yangpuensis DSM 45577 | Yangpumicins F, G | [88] |
| Nocardiopsis sp. LX-1 | Nocarpyrroline A, Isoflavonoid E | [89] |
| Nocardiopsis sp. SCA30 | 1-acetyl-4-4(hydroxyphenyl)piperazine | [90] |
| Pseudonocardia carboxydivorans M227 | Branimycins B, C | [91] |
| Streptomonospora sp. PA3 | Persiamycin A | [92] |
| Streptomyces aculeolatus PTM-029 | Napyradiomycins (3,7,9,10,12) | [93,94] |
| Streptomyces aculeolatus PTM-420 | Napyradiomycins (1,2,4–6,8,11) | [93] |
| Streptomyces albus 4N24 | Benzanthric acid | [95] |
| Streptomyces albus MAB56 | 12-methyltetradecanoic acid, Palmitic acid, Tridecanoic acid | [96] |
| Streptomyces armeniacus DSM 43125 | Armeniaspirol analogues 1–6, 9–12 | [94] |
| Streptomyces bacillaris MBTC38 | Lactoquinomycin A | [97] |
| Streptomyces californicus ADR1 | Methanoazulen-9-ol, decahydro-2, 2, 4, 8-tetramethyl-stereoisomer (Sesquiterpene) | [98] |
| Streptomyces coelicolor LY001 | 3-(3,5-dichloro-4-hydroxyphenyl) propanoic acid, 3-(3,5-dichloro-4-hydroxyphenyl) propanoic acid methyl ester, 3-(3-chloro-4-hydroxyphenyl) propanoic acid | [99] |
| Streptomyces diacarni LHW51701 | Chlocarbazomycins (CCBs) A–D | [100] |
| Streptomyces fradiae MM456M-mF7 | Fradiamines B | [101] |
| Streptomyces globisporus sp. WA5-2-37 | Actinomycin X2, Collismycin A | [102] |
| Streptomyces globisporus subsp. globisporus | Globimycin | [103] |
| Streptomyces griseoviridis PU-KB10–4 | Mitomycin C | [104] |
| Streptomyces hyaluromycini MB-PO13 | Rubromycins CA1, CA2 | [105] |
| Streptomyces koyangensis SCSIO 5802 | Neoabyssomicins F, G | [106] |
| Streptomyces malachitospinus ITD-35 | 3-octanone, neopentyl, Isothiocyanate, 2-methyl butyl isothiocyanate | [13] |
| Streptomyces microflavus (MBTI36) | Chromomycin A9, Ap, A2, A3 | [107] |
| Streptomyces lunaelactis MM109T | Lunaemycins A, B1, D | [108] |
| Streptomyces lusitanus OUCT16-27 | Grincamycin L | [109] |
| Streptomyces palmae CMU-AB204T | Phenyl alkenoic acids C, D Phenyl alkenoic acids E, F | [110] |
| Streptomyces qinglanensis 172205 | 15R-17,18-dehydroxantholipin | [111] |
| Streptomyces zhaozhouensis 208DD-064 | Streptopyrroles B, C | [112] |
| Streptomyces sp. 120454 | Mayamycin B | [113] |
| Streptomyces sp. 182SMLY | N-acetyl-N-demethylmayamycin | [114] |
| Streptomyces sp. 7NS3 | Emycin A | [115] |
| Streptomyces sp. 8P21H-1 | Desulphurzing, Griseoviridin, Griseoviridin | [116] |
| Streptomyces sp. AD-3-6 | Nybomycins D | [117] |
| Streptomyces sp. ADI91-18, ADI95-16 | Linearmycins | [117] |
| Streptomyces sp. ADI97-07 | Clavams | [117] |
| Streptomyces sp. ADI98-12 | Griselimycin, Kirromycin | [117] |
| Streptomyces sp. B9173 | Flaviogeranin B1, B, Flaviogeranin D, Flaviogeranin C2 | [118] |
| Streptomyces sp. CA-271078 | Napyradiomycins 2, D1 | [119] |
| Streptomyces sp. CPCC 204980 | Cervinomycins B1–B4 | [120] |
| Streptomyces sp. DSM14386 | Svetamycins C, G | [121] |
| Streptomyces sp. FJS31-2 | Zunyimycins B, C | [122] |
| Streptomyces sp. GKU 220 | Rakicidin F | [123] |
| Stretomyces sp. HCCB11876 | Quinomycins I, J | [124] |
| Streptomyces sp. HK-2006-1 | Aldgamycin O | [125] |
| Streptomyces sp. HN-A124 | Cysrabelomycin | [126] |
| Streptomyces sp. Hu103 | Anulamycins A-D | [127] |
| Streptomyces sp. HZP-2216E | 23-O-butyrylbafilomycin D | [128] |
| Streptomyces sp. IB201691-2A | Baikalomycins A–C | [129] |
| Streptomyces sp. ICN19 | Ala-geninthiocin | [130] |
| Streptomyces sp. KCB13F003 | Ulleungmycin A, B | [131] |
| Streptomyces sp. KCB14A132 | Enamidonins B, C | [132] |
| Streptomyces sp. MM168-141F8 | Quadoctomycin | [133] |
| Streptomyces sp. MNP32 | Phenol, 3,5-bis(1,1-dimethylethyl)-1,1′-Biphenyl]-2,3′-diol, 3,4′,5,6′-tetrakis(1,1-dimethylethyl) | [134] |
| Streptomyces sp. MUSC 125 | Thiophene, 2-butyl-5-ethyl, 1-Heptyn-3-ol, 8-[N-Aziridylethylamino]-2-6,dimethyloctene-2, Pyrrolo[1,2-a]pyrazine-1,4-dion,hexahydro, and 2,4-Dihydroxy-6-propylbenzoic acid | [42] |
| Streptomyces sp. NA06554 | Borrelidin J | [135] |
| Streptomyces sp. NA07423 | Nagimycins A, B | [136] |
| Streptomyces sp. OPOK_MB_B11 | Azalomycin F4a 2-ethylpentyl ester | [137] |
| Streptomyces sp. PBR11 | 1-Tetradecanol phenol, 2,5-bis(1,1-dimethylethyl n-pentadecanol, 1-nonadecene, and pyrrolo[1,2-a]pyrazine-1,4-dione hexahydro-3-(phenylmethyl). | [138] |
| Streptomyces sp. PNM-9 | 2-methyl-N-(2′-phenylethyl)-butanamide 3-methyl-N-(2′-phenylethyl)-butanamide | [139] |
| Streptomyces sp. SBT345 | Ageloline A | [140] |
| Streptomyces sp. SCSIO 41399 | Isotirandamycin B | [141] |
| Streptomyces sp. sima1_6 | Echinomycin | [137] |
| Streptomyces sp. SN0280 | Streptoone A | [142] |
| Streptomyces sp. SM01 | Picolinamycin | [143] |
| Streptomyces sp. SS | Sansanmycin Q | [144] |
| Streptomyces sp. Stup16_B49.2 | Echinomycin | [137] |
| Streptomyces sp. Stup16_B146 | TPU-0037-C | [137] |
| Streptomyces sp. Stup18_J70 | Bafilomycin A1 derivative | [137] |
| Streptomyces sp. UICC B-92 | 4-O-glucosyl,1-carboxyl-phenazine | [145] |
| Streptomyces sp. W367A | W367A | [137] |
| Streptomyces sp. XMA39 | Strepoxepinmycins A–D | [146] |
| Streptomyces sp. YINM00001 | Peperodione, Peperophthalene | [147] |
| Streptomyces sp. ZZ1118 | Streptoindoles A–D | [148] |
| Streptomyces sp. ZZ741 | Streptoglutarimides A–J | [149] |
| Streptomycete clade MAR4 CNY-960, CNS-284 | Marinocyanins A–F | [150] |
| Streptosporangium sp. SANK 60,501 | Muraminomicins A, B, C, D, E1, E2, F | [151] |
| Thermoactinomyces vulgaris ISCAR 2354 | Thermoactinoamide A | [152] |
| Verrucosispora sp. FIM06-0036 | 2-ethylhexyl 1H-imidazole-4-carboxylate | [100] |
| Verrucosispora sp. SCSIO 07399 | Kendomycins B–D | [153] |
* Most records were published in the PubMed and Web of Science databases from 2016 to 2023.
Table 2.
Examples of antifungal metabolites identified from Actinomycetota from 2016 to 2023 *.
| Organism (s) | Compound Name (s) | Reference (s) |
|---|---|---|
| Actinokineospora bangkokensis 44EHWT | Thailandins A & B | [159] |
| Actinomadura sp. BCC 35,430 | Actinomadurone | [161] |
| Amycolatopsis sp. M39 | Macrotermycins A & D | [70] |
| Acrocarpospora punica 04107M | Acrocarposporins A, B, D, and E | [162] |
| Dermabacter vaginalis AD1-86 | Dermazolium A | [163] |
| Micromonospora sp. UR56 + Actinokineospora sp. EG49 | Phenazine-1-carboxylic acid, Aestivophoenin c, Methyl saphenate | [80] |
| Nocardia abscessus IFM 10029 | Nabscessins A, B | [164] |
| Nocardiopsis sp. LX-1 | Isoflavonoid E | [89] |
| Pseudonocardia autotrophica KCTC9441 | Polyene B1 | [165] |
| Saccharothrix yanglingensis Hhs.015 | 10-deoxyfungichromin (WH02) | [166] |
| Streptomyces albidoflavus STV1572a | 1-heneicosanol | [167] |
| Streptomyces albolongus YIM 101047 | 19-methoxybafilomycin C1 amide | [168] |
| Streptomyces antibioticus strain 200-09 | Kitamycin C | [169] |
| Streptomyces caniferus GUA-06-05-006A | PM100117, PM100118 | [170] |
| Streptomyces griseoviridis PU-KB10–4 | Mitomycin C | [104] |
| Streptomyces hyaluromycini MB-PO13 | Rubromycins CA1, CA2 | [105] |
| Streptomyces morookaense AM25 | Gloeosporiocide | [171] |
| Streptomyces sp. 1H-XA2 | Furamycins I, II | [172] |
| Streptomyces sp. ADI91-18 | Linearmycins | [117] |
| Streptomyces sp. ADI95-16 | Linearmycins | [117] |
| Streptomyces sp. ADI96-02 | Cycloheximide, Galbonolides | [117] |
| Streptomyces sp. ADI96-15 | Candicidin | [117] |
| Streptomyces sp. ADI97-07 | Galbonolides | [117] |
| Streptomyces sp. ADI98-12 | Candicidin | [117] |
| Streptomyces sp. BV410 | Staurosporine | [173] |
| Streptomyces sp. CB09030 | LOB A, B, H8 | [174] |
| Streptomyces sp. CT37 | Legonimide 1, 1H-indole-3-carbaldehyde | [175] |
| Streptomyces sp. FX13 | Oligomycin A | [176] |
| Streptomyces sp. HAAG3-15 | Azalomycin B | [177] |
| Streptomyces sp. KIB-H869 | Hygrolidin-type macrolide | [178] |
| Streptomyces sp. MUSC 125 | Thiophene, 2-butyl-5-ethyl, 1-Heptyn-3-ol, Pyrrolo[1,2-a] pyrazine-1,4-dion,hexahydro, 9,9-Dimethyl-3, 7-diazabicyclo[3.3.1]nonane, 2,4-Dihydroxy-6-propylbenzoic acid | [42] |
| Streptomyces sp. PBR11 | 1-tetradecanol, n-pentadecanol, 1-nonadecene, pyrrolo [1,2-a]pyrazine-1,4-dione hexahydro-3-(phenylmethyl) | [138] |
| Streptomyces sp. QHH-9511 | 6-deoxy-13-hydroxy-8,11-dione Dihydrogranaticin A Granaticin A, B | [179] |
| Streptomyces sp. SN0280 | Streptoone B | [142] |
| Streptomyces sp. SY1965 | Streptothiazomycin A, Streptodiketopiperazines A, B, [2-hydroxy-1-(hydroxymethyl)ethyl]-2-methoxybenzamide, salicylamide, 4-hydroxymethyl benzoate, Spoxazomicin C | [180] |
| Streptomyces sp. TR1341 | Filipin, Fungichromin, Actinomycin X2 | [181] |
| Streptomyces sp. TT3 | Actinorhodin | [182] |
| Streptomyces sp. YO15-A001 | YO-001A | [183] |
| Streptomyces albus CAI-21 | Organophosphate | [184] |
| Streptomyces coelicolor LY001 | diketopiperazine alkaloids cyclo (l-Phe-trans-4-OH-l-Pro), cyclo(l-Phe-cis-4-OH-d-Pro) | [99] |
| Streptomyces palmae CMU-AB204T | Phenyl alkenoic acids A,B, Anguinomycin A, Leptomycin A, Actinopyrone A | [110] |
| Streptomyces qinglanensis 172205 | 15R-17,18-dehydroxantholipin | [111] |
| Streptomycete clade MAR4 CNY-960 and CNS-284 | Marinocyanins A | [150] |
| Umezawaea sp. RD066910 + Tsukamurella pulmonis TP-B0596 | Umezawamides A | [185] |
* Most records were published in the PubMed and Web of Science databases from 2016 to 2023.
Table 4.
Examples of antitumor metabolites identified from Actinomycetota from 2016 to 2023 *.
| Organism (s) | Compound Name (s) | Reference (s) |
|---|---|---|
| Actinoalloteichus hymeniacidonis 179DD-027 | Dokdolipid B | [198] |
| Actinomadura sp. K13-0306 | Sagamilactam | [199] |
| Actinomadura sp. K4S16 | Nonthmicin, Ecteinamycin | [200] |
| Actinosynnema pretiosum HGF052::asm18 | Actinosynneptide A, B | [201] |
| Amycolatopsis sp. IRD-009 | Pradimicin-IRD | [69] |
| Amycolatopsis alba DSM 44,262 | Thioalbamide | [202] |
| Catenuloplanes sp. RD067331 + Tsukamurella pulmonis TP-B059 | Catenulobactin B | [203] |
| Micromonospora sp. UR56 + Actinokineospora sp. EG49 | Phenazine-1-carboxylic acid, Aestivophoenin c, Methyl saphenate | [80,204] |
| Micromonospora carbonacea LS276 | Tetrocarcin Q | [76] |
| Micromonospora zhangzhouensis HM134T | 7E,11E)-6-hydroxy-1-isopropyl-11-(methoxycarbonyl)-4-methylene-1,2,3,4,4a,5,6,9,10,12a-decahydrobenzo[10]annulene-7-carboxylic acid | [205] |
| Micromonospora matsumotoense M-412 | Paulomycin G | [206] |
| Micromonospora aurantiaca 110B | Isoflavonoid glycosides 1-3 | [207] |
| Micromonospora chalcea FIM 02-523 | Rakicidins G–I | [208] |
| Micromonospora sp. FIM05328 | FW05328-1 | [209] |
| Micromonospora sp. HS-HM-036 | Naphthalenepropanoic acid analog | [210] |
| Micromonospora yangpuensis DSM 45577 | Yangpumicin A | [211] |
| Micromonospora yangpuensis DSM 45577 | Yangpumicins F, G | [88] |
| Nocardiopsis alba KM6-1 | Isomethoxyneihumicin | [212] |
| Nocardiopsis sp. CG3 | Kenalactams A, E | [213] |
| Nonomuraea endophytica GW58/450 | Karamomycins B, C | [214] |
| Nonomuraea sp. AKA32 | Akazamicin | [215] |
| Saccharomonospora sp. UR22 + Dietzia sp. UR66 | Saccharomonosporine A | [216] |
| Streptomonospora sp. PA3 | Persiamycin A | [92] |
| Streptomyces albolongus YIM 101047 | 19-methoxybafilomycin C1 amide, 21-deoxybafilomycin A1 | [168] |
| Streptomyces ardesiacus 156VN-095 | Urdamycins W, X, Grincamycin U | [217] |
| Streptomyces bacillaris | 2,4-di-tert-butylphenol | [218] |
| Streptomycete clade MAR4 CNY-960 and CNS-284 | Marinocyanins A–F | [150] |
| Streptomyces qinglanensis 172205 | 15R-17,18-dehydroxantholipin, (3E,5E,7E)-3-methyldeca-3,5,7-triene-2,9-dione, Qinlactone A–B | [111] |
| Streptomyces cacaoi subsp. asoensis H2S5 | Trienomycins J–L | [219] |
| Streptomyces caniferus GUA-06-05-006A | PM100117, PM100118 | [220] |
| Streptomyces coelicolor M1146 | Staurosporines M1, M2 | [221] |
| Streptomyces curacoi NBRC 12761 | Curacozole | [222] |
| Streptomyces cyaneofuscatus M-157 | 3-Hydroxyquinaldic acid derivative 1 | [223] |
| Streptomyces griseoviridis 2464-S5 | Prodigiosin R2 | [224] |
| Streptomyces seoulensis A01 | Ansaseomycins A, B | [225] |
| Streptomyces subflavus subsp. irumaensis AM-3603 | Bisoxazolomycin A | [226] |
| Streptomyces violascens YIM 100225 | Albaflavenoid | [227] |
| Streptomyces violascens YIM100212 | Fusicomycin A, B, Isofusicomycin A | [228] |
| Streptomyces sp. 166 | Sekgranaticin | [229] |
| Streptomyces sp. 182SMLY | N-acetyl-N-demethylmayamycin, Streptoanthraquinone A | [230] |
| Streptomyces sp. 1H-GS5 | Spectinabilin derivative 1 | [231] |
| Streptomyces sp. A65 | Streptocarbazoles C, 2040-epi-K252d, 0-epi-K252d | [232] |
| Streptomyces sp. AD-3 | Nybomycins D | [233] |
| Streptomyces sp. ADI92-24 | Tomaymycin | [117] |
| Streptomyces sp. ADI95-17 | Naringenin | [117] |
| Streptomyces sp. ADI96-02 | Echinomycin | [117] |
| Streptomyces sp. ADI96-15 | Antimycin, Lobophorins | [117] |
| Streptomyces sp. ADI97-07 | Neocarzilin, Antimycin | [117] |
| Streptomyces sp. ADI98-10 | Actinomycin | [117] |
| Streptomyces sp. B9173 | Flaviogeranin B1, B, C2, D | [118] |
| Streptomyces sp. BB47 | Antarlides F–H | [234] |
| Streptomyces sp. CHQ-64 ΔrdmF | Geranylpyrrol A, Piericidin F | [235] |
| Streptomyces sp. CMAA 1527 | Cinerubin B | [15] |
| Streptomyces sp. CMAA 1653 | Actinomycin V | [15,120] |
| Streptomyces sp. CPCC 204,980 | Cervinomycins B1-B4 | [120] |
| Streptomyces sp. DT-A61 | 9-hydroxy-K252c, 3-hydroxy-K252c, 3-hydroxy-7-methoxy-K252c, Nacetylholyrine A, 3-hydroxyholyrine A, 3′-O-demethyl-4′-N-demethyl-4′-N-acetyl-4′-epi-staurosporine, Streptocarbazoles D, E | [236] |
| Streptomyces sp. EGY1 | Sharkquinone | [237] |
| Streptomyces sp. GIC10-1 | Bafilomycin M | [238] |
| Streptomyces sp. GKU 220 | Rakicidin F | [123] |
| Streptomyces sp. HF-11225 | Nivelactam B | [239] |
| Streptomyces sp. HN-A124 | Cysrabelomycin | [126] |
| Streptomyces sp. HNA39 | Cyclizidines B–I | [240] |
| Streptomyces sp. HS-NF-1178 | 218-seco-lankacidinols A, B | [241] |
| Streptomyces sp. HS-NF-780 | 9-methylstreptimidone 2-α-d-glucopyranoside, ydroxyiso-9-methylstreptimidone | [242] |
| Streptomyces sp. HZP-2216E | 23-O-butyrylbafilomycin D | [81,128] |
| Streptomyces sp. ICN19 | Ala-geninthiocin | [130] |
| Streptomyces sp. IFM 11490 | Elmenols G | [243] |
| Streptomyces sp. KCB13F003 | Ulleungdin | [244] |
| Streptomyces sp. KCB13F030 | Ulleungoside | [245] |
| Streptomyces sp. KIB-H1318 | Phenoxazinone-related alkaloid 2 | [246] |
| Streptomyces sp. KIB-H714 | Actinomycin Z6 | [247] |
| Streptomyces sp. M268 | Kiamycin B | [248] |
| Streptomyces sp. MSB090213SC12 | Neothioviridamide | [249] |
| Streptomyces sp. MUSC 125 | 8-[N-Aziridylethylamino]-2-6,dimethyloctene-2 | [42] |
| Streptomyces sp. N1510.2 | Strepantibin D | [250] |
| Streptomyces sp. N1510.2 | Strepantibins A–C | [251] |
| Streptomyces sp. NA4286 | Murayaquinone D | [252] |
| Streptomyces sp. NEAU-L3 | Tetracenoquinocin A | [253] |
| Streptomyces sp. OPMA00071 | JBIR-150 | [254] |
| Streptomyces sp. PU-14G | Puromycins C, D | [255] |
| Streptomyces sp. PU-KB10-4 | 4-hydroxycinnamide | [104] |
| Streptomyces sp. Q22 | Bagremycins C | [256] |
| Streptomyces sp. RAI364 | Curromycin A | [257] |
| Streptomyces sp. RK88 | Opantimycin A | [258] |
| Streptomyces sp. SBT345 | Strepoxazine A | [259] |
| Streptomyces sp. SCSIO 03032 | Indimicins F, G, Spiroindimicins G, H | [260] |
| Streptomyces sp. SCSIO 1666/17C4 (engineered) | l-rhodinose-l-rhodinose-2-deoxy-l-fucose-10-decarbomethoxy-ε-rhodomycinone | [219] |
| Streptomyces sp. SCSIO 40063 | Piericidins A5, G1 | [261] |
| Streptomyces sp. SCSIO 41399 | Aranciamycin K, Isotirandamycin B | [141] |
| Streptomyces sp. SS13I | Ephyromycins B, C | [262] |
| Streptomyces sp. SS17F | Thioquinomycins A, C, D | [263] |
| Streptomyces sp. SSA28 | Cytosaminomycin E | [264] |
| Streptomyces sp. TR1341 | Filipin, Fungichromin, Actinomycin X2 | [181] |
| Streptomyces sp. XMA39 | Strepoxepinmycins A–D | [146] |
| Streptomyces sp. W2061 | Rubiflavin G Photorubiflavin G, E | [265] |
| Streptomyces sp. YIM S01863 | Jiangchuanmycin | [266] |
| Streptomyces sp. ZZ741 | Streptoglutarimides H | [149] |
| Streptomyces OPOK_MB_A9 | Milbemycin-α8 | [137] |
| Streptomyces sp. shell-016 | Shellmycin A–D | [267] |
| Umezawaea sp. RD066910 + Tsukamurella pulmonis TP-B0596 | Umezawamides A & B | [185] |
| Verrucosispora sp. SCSIO 07399 | Kendomycins B-D | [153] |
* Most records were published in the PubMed and Web of Science databases from 2016 to 2023.
Table 5.
Examples of antiviral metabolites identified from Actinomycetota from 2016 to 2023 *.
| Organism (s) | Compound Name (s) | Reference (s) |
|---|---|---|
| Actinomadura sp. 2EPS | Decatromicins | [278] |
| Kibdelosporangium persicum | Persicamidines A–E | [279] |
| Kutzneria albida DSM 43870 | Huimycin | [106,280] |
| Streptomyces kebangsaanensis WS-68302 | Napyradiomycin A4, A80915 H | [281] |
| Streptomyces jiujiangensis NBERC-24992 | Virantmycins D–G | [282] |
| Streptomyces bacillaris | Zelkovamycins F, G | [283] |
| Streptomyces koyangensis SCSIO 5802 | Neoabyssomicins F, G | [106,284] |
| Streptomyces sp. AM-2504 | Virantmycins B | [284,285] |
| Streptomyces sp. CPCC 200267 | Geninthiocins E, F | [286] |
| Streptomyces sp. HK18 | Xiamycins D | [285,287] |
| Streptomyces sp. JA74 | Dihydromaniwamycin E | [288] |
| Streptomyces sp. SMU 03 | dichloromethane extracts (DCME) | [287] |
* Most records were published in the PubMed and Web of Science databases from 2016 to 2023.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ngamcharungchit, C.; Chaimusik, N.; Panbangred, W.; Euanorasetr, J.; Intra, B. Bioactive Metabolites from Terrestrial and Marine Actinomycetes. Molecules 2023, 28, 5915. https://doi.org/10.3390/molecules28155915
AMA Style
Ngamcharungchit C, Chaimusik N, Panbangred W, Euanorasetr J, Intra B. Bioactive Metabolites from Terrestrial and Marine Actinomycetes. Molecules. 2023; 28(15):5915. https://doi.org/10.3390/molecules28155915
Chicago/Turabian StyleNgamcharungchit, Chananan, Nutsuda Chaimusik, Watanalai Panbangred, Jirayut Euanorasetr, and Bungonsiri Intra. 2023. "Bioactive Metabolites from Terrestrial and Marine Actinomycetes" Molecules 28, no. 15: 5915. https://doi.org/10.3390/molecules28155915