
Cell Cycle, Telomeres, and Telomerase in Leishmania spp.: What Do We Know So Far?

 , , , ,
, , , ,  and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
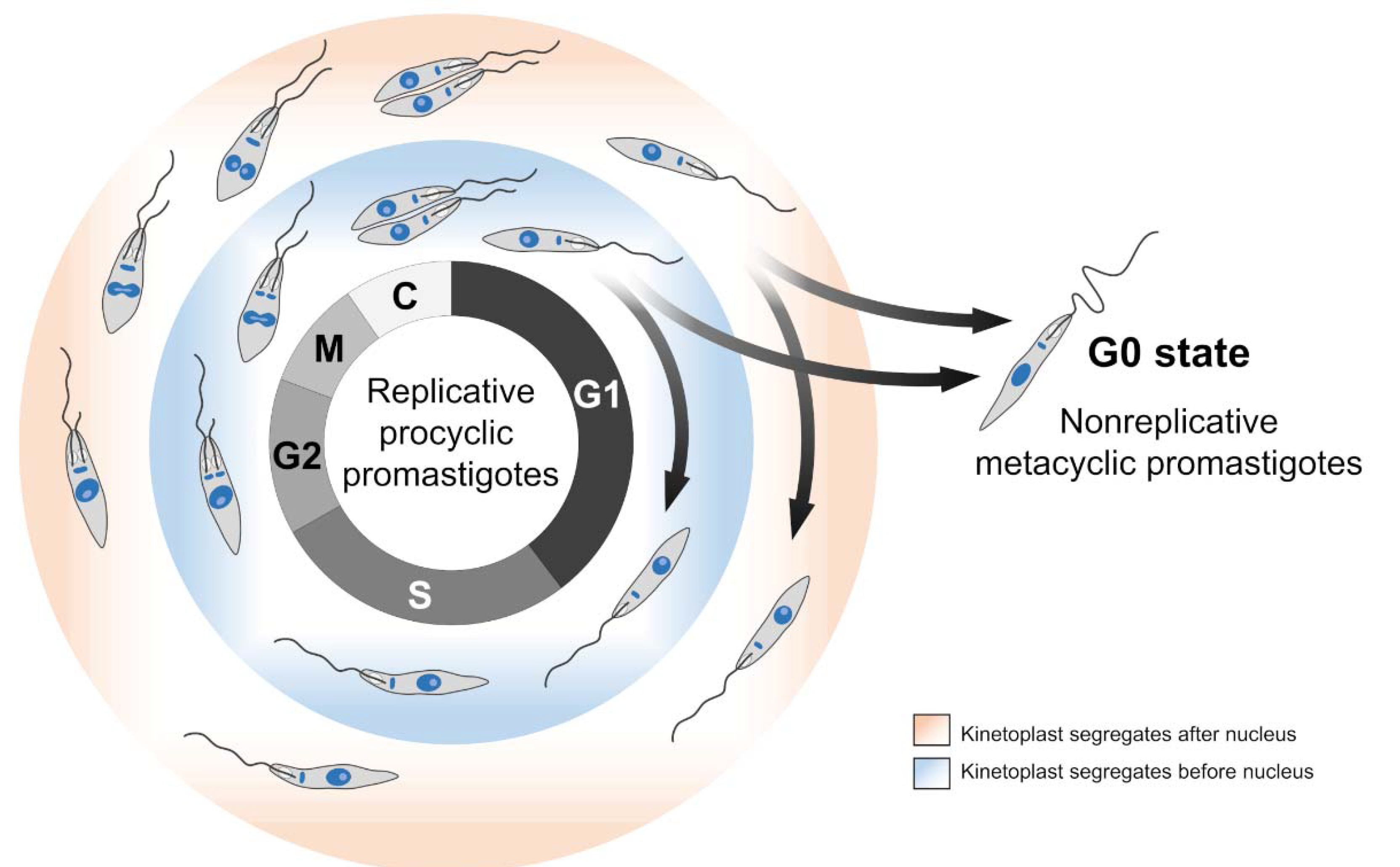
2. Leishmania spp. Cell Cycle
3. Leishmania spp. Telomeres
4. Leishmania spp. Telomerase
4.1. Structure and Function
4.2. Biogenesis and Mechanisms
4.3. Phylogenetic Context of Leishmania spp. Telomerases among Other Pathogenic Trypanosomatids
5. Conclusions Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cantanhêde, L.M.; Mata-Somarribas, C.; Chourabi, K.; Pereira da Silva, G.; Dias das Chagas, B.; de Oliveira, R.; Pereira, L.; Côrtes Boité, M.; Cupolillo, E. The Maze Pathway of Coevolution: A Critical Review over the Leishmania and Its Endosymbiotic History. Genes 2021, 12, 657. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Serafim, T.D.; Coutinho-Abreu, I.V.; Dey, R.; Kissinger, R.; Valenzuela, J.G.; Oliveira, F.; Kamhawi, S. Leishmaniasis: The Act of Transmission. Trends Parasitol. 2021, 37, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Farhoudi, R. Leishmaniasis in Humans: Drug or Vaccine Therapy? Drug Des. Dev. Ther. 2017, 12, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Cruz, A.K.; Freitas-Castro, F. Genome and Transcriptome Analyses of Leishmania spp.: Opening Pandora’s Box. Curr. Opin. Microbiol. 2019, 52, 64–69. [Google Scholar] [CrossRef]
- Lander, N.; Chiurillo, M.A. State-of-the-art CRISPR/Cas9 Technology for Genome Editing in Trypanosomatids. J. Eukaryot. Microbiol. 2019, 66, 981–991. [Google Scholar] [CrossRef]
- Vasconcelos, E.J.R.; Nunes, V.S.; da Silva, M.S.; Segatto, M.; Myler, P.J.; Cano, M.I.N. The Putative Leishmania Telomerase RNA (LeishTER) Undergoes Trans-Splicing and Contains a Conserved Template Sequence. PLoS ONE 2014, 9, e112061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandin, S.; Rhodes, D. Telomerase Structure. Curr. Opin. Struct. Biol. 2014, 25, 104–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podlevsky, J.D.; Chen, J.J.-L. Evolutionary Perspectives of Telomerase RNA Structure and Function. RNA Biol. 2016, 13, 720–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Espinal, A.; Cross, G.A.M. Trypanosome Telomeres Are Protected by a Homologue of Mammalian TRF2. Mol. Cell. Biol. 2005, 25, 5011–5021. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, C.A.H.; Perez, A.M.; Barros, A.C.; Dreyer, T.R.; da Silva, M.S.; Morea, E.G.O.; Fontes, M.R.M.; Cano, M.I.N. Dual Cellular Localization of the Leishmania Amazonensis Rbp38 (LaRbp38) Explains Its Affinity for Telomeric and Mitochondrial DNA. Biochimie 2019, 162, 15–25. [Google Scholar] [CrossRef]
- Fernandes, C.A.H.; Morea, E.G.O.; dos Santos, G.A.; da Silva, V.L.; Vieira, M.R.; Viviescas, M.A.; Chatain, J.; Vadel, A.; Saintomé, C.; Fontes, M.R.M.; et al. A Multi-Approach Analysis Highlights the Relevance of RPA-1 as a Telomere End-Binding Protein (TEBP) in Leishmania Amazonensis. Biochim. Biophys. Acta BBA-Gen. Subj. 2020, 1864, 129607. [Google Scholar] [CrossRef]
- Dey, A.; Chakrabarti, K. Current Perspectives of Telomerase Structure and Function in Eukaryotes with Emerging Views on Telomerase in Human Parasites. Int. J. Mol. Sci. 2018, 19, 333. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.G.; Pouchkina-Stantcheva, N.; Di Donfrancesco, A.; Kildisiute, G.; Sahu, S.; Aboobaker, A.A. The Protein Subunit of Telomerase Displays Patterns of Dynamic Evolution and Conservation across Different Metazoan Taxa. BMC Evol. Biol. 2017, 17, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, B.C.D.; Shiburah, M.E.; Paiva, S.C.; Vieira, M.R.; Morea, E.G.O.; da Silva, M.S.; Alves, C.S.; Segatto, M.; Gutierrez-Rodrigues, F.; Borges, J.C.; et al. Possible Involvement of Hsp90 in the Regulation of Telomere Length and Telomerase Activity During Leishmania amazonensis Developmental Cycle and Population Proliferation. Front. Cell Dev. Biol. 2021, in press. [Google Scholar] [CrossRef]
- Cano, M.I.N. Telomere Biology of Trypanosomatids: More Questions than Answers. Trends Parasitol. 2001, 17, 425–429. [Google Scholar] [CrossRef]
- Lira, C.B.B.; Giardini, M.A.; Neto, J.L.S.; Conte, F.F.; Cano, M.I.N. Telomere Biology of Trypanosomatids: Beginning to Answer Some Questions. Trends Parasitol. 2007, 23, 357–362. [Google Scholar] [CrossRef]
- Damasceno, J.D.; Silva, G.L.; Tschudi, C.; Tosi, L.R. Evidence for Regulated Expression of Telomeric Repeat-Containing RNAs (TERRA) in Parasitic Trypanosomatids. Mem. Inst. Oswaldo Cruz 2017, 112, 572–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poláková, E.; Záhonová, K.; Albanaz, A.T.S.; Butenko, A.; Lukeš, J.; Yurchenko, V. Diverse Telomeres in Trypanosomatids. Parasitology 2021, 148, 1254–1270. [Google Scholar] [CrossRef]
- Blackburn, E.H. Telomeres: Structure and Synthesis. J. Biol. Chem. 1990, 265, 5919–5921. [Google Scholar] [CrossRef]
- Turner, K.; Vasu, V.; Griffin, D. Telomere Biology and Human Phenotype. Cells 2019, 8, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greider, C.W. Telomere Length Regulation. Annu. Rev. Biochem. 1996, 65, 337–365. [Google Scholar] [CrossRef]
- Barbé-Tuana, F.; Grun, L.K.; Pierdoná, V.; de Oliveira, B.C.D.; Paiva, S.C.; Shiburah, M.E.; da Silva, V.L.; Morea, E.G.O.; Fontes, V.S.; Cano, M.I.N. Human Chromosome Telomeres. In Human Genome Structure, Function and Clinical Considerations; Haddad, L.A., Ed.; Springer: Cham, Switzerland, 2021; pp. 207–243. ISBN 978-3-030-73150-2. [Google Scholar]
- Pierce, B.A. Genetics: A Conceptual Approach; Editora Guanabara Koogan: Rio de Janeiro, Brazil, 2016; v. 5. [Google Scholar]
- Greider, C.W. Regulating Telomere Length from the Inside Out: The Replication Fork Model. Genes Dev. 2016, 30, 1483–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, W.E.; Tesmer, V.M.; Huffman, K.E.; Levene, S.D.; Shay, J.W. Normal Human Chromosomes Have Long G-Rich Telomeric Overhangs at One End. Genes Dev. 1997, 11, 2801–2809. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.D. Origin of Concatemeric T7DNA. Nat. New Biol. 1972, 239, 197–201. [Google Scholar] [CrossRef]
- Hayflick, L. The Limited in vitro Lifetime of Human Diploid Cell Strains. Exp. Cell Res. 1965, 37, 614–636. [Google Scholar] [CrossRef]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres Shorten during Ageing of Human Fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, L.; Wang, Z.; Liu, J.-P. Roles of Telomere Biology in Cell Senescence, Replicative and Chronological Ageing. Cells 2019, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Greider, C.W.; Blackburn, E.H. The Telomere Terminal Transferase of Tetrahymena Is a Ribonucleoprotein Enzyme with Two Kinds of Primer Specificity. Cell 1987, 51, 887–898. [Google Scholar] [CrossRef]
- Giardini, M.A.; Segatto, M.; da Silva, M.S.; Nunes, V.S.; Cano, M.I.N. Telomere and Telomerase Biology. In Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2014; Volume 125, pp. 1–40. [Google Scholar] [CrossRef]
- Berei, J.; Eckburg, A.; Miliavski, E.; Anderson, A.D.; Miller, R.; Dein, J.; Giuffre, A.M.; Tang, D.; Deb, S.; Racherla, K.S.; et al. Potential Telomere-related Pharmacological Targets. Curr. Top. Med. Chem. 2019, 20, 458–484. [Google Scholar] [CrossRef]
- Terzi, M.Y.; Izmirli, M.; Gogebakan, B. The Cell Fate: Senescence or Quiescence. Mol. Biol. Rep. 2016, 43, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Gossage, S.M.; Rogers, M.E.; Bates, P.A. Two Separate Growth Phases during the Development of Leishmania in Sand Flies: Implications for understanding the life cycle. Int. J. Parasitol. 2003, 33, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Rittershaus, E.S.C.; Baek, S.H.; Sassetti, C.M. The Normalcy of Dormancy: Common Themes in Microbial Quiescence. Cell Host Microbe 2013, 13, 643–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, R.J.; Gluenz, E.; Gull, K. The Cell Cycle of Leishmania: Morphogenetic Events and Their Implications for Parasite Biology. Mol. Microbiol. 2011, 79, 647–662. [Google Scholar] [CrossRef] [Green Version]
- Ambit, A.; Woods, K.L.; Cull, B.; Coombs, G.H.; Mottram, J.C. Morphological Events during the Cell Cycle of Leishmania major. Eukaryot. Cell 2011, 10, 1429–1438. [Google Scholar] [CrossRef] [Green Version]
- Simpson, L.; Braly, P. Synchronization of Leishmania tarentolae by Hydroxyurea. J. Protozool. 1970, 17, 511–517. [Google Scholar] [CrossRef]
- da Silva, M.S.; Monteiro, J.P.; Nunes, V.S.; Vasconcelos, E.J.; Perez, A.M.; Freitas-Júnior, L.H.; Elias, M.C.; Cano, M.I.N. Leishmania amazonensis Promastigotes Present Two Distinct Modes of Nucleus and Kinetoplast Segregation during Cell Cycle. PLoS ONE 2013, 8, e81397. [Google Scholar] [CrossRef] [Green Version]
- Minocha, N.; Kumar, D.; Rajanala, K.; Saha, S. Kinetoplast morphology and segregation pattern as a marker for cell cycle progression in Leishmania donovani. J. Eukaryot. Microbiol. 2011, 58, 249–253. [Google Scholar] [CrossRef]
- da Silva, M.S.; Muñoz, P.A.M.; Armelin, H.A.; Elias, M.C. Differences in The Detection of BrdU/EdU Incorporation Assays Alter the Calculation for G1, S, and G2 Phases of the Cell Cycle in Trypanosomatids. J. Eukaryot. Microbiol. 2017, 64, 756–770. [Google Scholar] [CrossRef]
- Archer, S.K.; Inchaustegui, D.; Queiroz, R.; Clayton, C. The Cell Cycle Regulated Transcriptome of Trypanosoma brucei. PLoS ONE 2011, 6, e18425. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.S.; Cayres-Silva, G.R.; Vitarelli, M.O.; Marin, P.A.; Hiraiwa, P.M.; Araújo, C.B.; Scholl, B.B.; Ávila, A.R.; McCulloch, R.; Reis, M.S.; et al. Transcription Activity Contributes to The Firing of Non-constitutive Origins in African trypanosomes Helping to Maintain Robustness in S-phase Duration. Sci. Rep. 2019, 9, 18512. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.S.; Pavani, R.S.; Damasceno, J.D.; Marques, C.A.; McCulloch, R.; Tosi, L.R.O.; Elias, M.C. Nuclear DNA Replication in Trypanosomatids: There Are No Easy Methods for Solving Difficult Problems. Trends Parasitol. 2017, 33, 858–874. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.A.; Dickens, N.J.; Paape, D.; Campbell, S.J.; McCulloch, R. Genome-wide Mapping Reveals Single-origin Chromosome Replication in Leishmania, a Eukaryotic Microbe. Genome Biol. 2015, 16, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, M.S.; Vitarelli, M.O.; Souza, B.F.; Elias, M.C. Comparative Analysis of The Minimum Number of Replication Origins in Trypanosomatids And Yeasts. Genes 2020, 11, 523. [Google Scholar] [CrossRef] [PubMed]
- Harashima, H.; Dissmeyer, N.; Schnittger, A. Cell Cycle Control across the Eukaryotic Kingdom. Trends Cell Biol. 2013, 23, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Liu, Y.; Zhou, Q.; Siegel, S.; Li, Z. The Centriole Cartwheel Protein SAS-6 in Trypanosoma brucei Is Required for Probasal Body Biogenesis and Flagellum Assembly. Eukaryot. Cell 2015, 14, 898–907. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, R.J.; Sunter, J.D.; Gull, K. Flagellar Pocket Restructuring through the Leishmania Life Cycle Involves a Discrete Flagellum Attachment zone. J. Cell Sci. 2016, 129, 854–867. [Google Scholar] [CrossRef] [Green Version]
- Lodish, H.; Berk, A. Overview of the Cell Cycle, and Its Control. In Molecular Cell Biology, 4th ed.; W. H. Freeman: New York, NY, USA, 2000; Section 13.1. [Google Scholar]
- Campbell, P.C.; De Graffenried, C.L. Alternate Histories of Cytokinesis: Lessons from the Trypanosomatids. Mol. Biol. Cell 2020, 31, 2631–2639. [Google Scholar] [CrossRef]
- Hecker, H.; Gander, E.S. The Compaction Pattern of the Chromatin of Trypanosomes. Biol. Cell 1985, 53, 199–208. [Google Scholar] [CrossRef]
- Hecker, H.; Betschart, B.; Bender, K.; Burri, M.; Schlimme, W. The Chromatin of Trypanosomes. Int. J. Parasitol. 1994, 24, 809–819. [Google Scholar] [CrossRef]
- Wheeler, R.J.; Gluenz, E.; Gull, K. The Limits on Trypanosomatid Morphological Diversity. PLoS ONE 2013, 8, e79581. [Google Scholar] [CrossRef] [Green Version]
- Shlomai, J. The Structure and Replication of Kinetoplast DNA. Curr. Mol. Med. 2005, 4, 623–647. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Y.; Motyka, S.A.; Agbo, E.E.C.; Englund, P.T. Fellowship of the Rings: The Replication of Kinetoplast DNA. Trends Parasitol. 2005, 21, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Valdivia, H.O.; Reis-Cunha, J.L.; Rodrigues-Luiz, G.F.; Baptista, R.P.; Baldeviano, G.C.; Gerbasi, R.V.; Dobson, D.E.; Pratlong, F.; Bastien, P.; Lescano, A.G.; et al. Comparative Genomic Analysis of Leishmania (Viannia) Peruviana and Leishmania (Viannia) braziliensis. BMC Genom. 2015, 16, 715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiurillo, M.A.; Beck, A.E.; Devos, T.; Myler, P.J.; Stuart, K.; Ramirez, J.L. Cloning and Characterization of Leishmania Donovani Telomeres. Exp. Parasitol. 2000, 94, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Baker, D.C. Characterisation of Leishmania Telomeres Reveals Unusual Telomeric Repeats and Conserved Telomere-Associated Sequence. Nucleic Acids Res. 1998, 26, 2161–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conte, F.F.; Cano, M.I.N. Genomic Organization of Telomeric and Subtelomeric Sequences of Leishmania (Leishmania) Amazonensis. Int. J. Parasitol. 2005, 35, 1435–1443. [Google Scholar] [CrossRef]
- Genest, P.A.; Borst, P. Analysis of Telomere Length Variation in Leishmania over Time. Mol. Biochem. Parasitol. 2007, 151, 213–215. [Google Scholar] [CrossRef]
- Pryde, F.E.; Gorham, H.C.; Louis, E.J. Chromosome ends: All the same under their caps. Curr. Opin. Genet. Dev. 1997, 7, 822–828. [Google Scholar] [CrossRef]
- Ravel, C.; Wincker, P.; Bastien, P.; Blaineau, C.; Pagès, M. A polymorphic minisatellite sequence in the subtelomeric regions of chromosomes I and V in Leishmania infantum. Mol. Biochem. Parasitol. 1995, 74, 31–41. [Google Scholar] [CrossRef]
- Dmitriev, P.V.; Petrov, A.V.; Dontsova, O.A. Yeast Telosome Complex: Components and Their Functions. Biochem. Mosc. 2003, 68, 718–734. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.S.; Silveira, R.C.V.; Perez, A.M.; Monteiro, J.P.; Calderano, S.G.; Da Cunha, J.P.; Elias, M.C.; Cano, M.I.N. Nuclear DNA Replication in Trypanosomatid Protozoa. In DNA Replication and Mutation; Leitner, R., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2012. [Google Scholar]
- Lira, C.B.B.; de Siqueira Neto, J.L.; Khater, L.; Cagliari, T.C.; Peroni, L.A.; dos Reis, J.R.R.; Ramos, C.H.I.; Cano, M.I.N. LaTBP1: A Leishmania Amazonensis DNA-Binding Protein That Associates in vivo with Telomeres and GT-Rich DNA Using a Myb-like Domain. Arch. Biochem. Biophys. 2007, 465, 399–409. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.S.; Perez, A.M.; Silveira, R.C.; Moraes, C.E.; Siqueira-Neto, J.L.; Freitas-Junior, L.H.; Cano, M.I.N. The Leishmania amazonensis TRF (TTAGGG Repeat Binding Factor) Homologue Binds and Co-localizes with Telomeres. BMC Microbiol. 2010, 10, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, M.F.; Castellari, R.R.; Conte, F.F.; Gozzo, F.C.; Sabino, A.A.; Pinheiro, H.; Novello, J.C.; Eberlin, M.N.; Cano, M.I.N. Identification of Three Proteins That Associate in vitro with The Leishmania (Leishmania) amazonensis G-rich Telomeric Strand: G-telomeric Proteins in L. amazonensis. Eur. J. Biochem. 2004, 271, 3050–3063. [Google Scholar] [CrossRef]
- Lira, C.B.B.; Siqueira Neto, J.L.; Giardini, M.A.; Winck, F.V.; Ramos, C.H.I.; Cano, M.I.N. LaRbp38: A Leishmania Amazonensis Protein That Binds Nuclear and Kinetoplast DNAs. Biochem. Biophys. Res. Commun. 2007, 358, 854–860. [Google Scholar] [CrossRef]
- Siqueira-Neto, J.L.; Lira, C.B.B.; Giardini, M.A.; Khater, L.; Perez, A.M.; Peroni, L.A.; dos Reis, J.R.R.; Freitas-Junior, L.H.; Ramos, C.H.I.; Cano, M.I.N. Leishmania Replication Protein A-1 Binds in vivo Single-stranded Telomeric DNA. Biochem. Biophys. Res. Commun. 2007, 358, 417–423. [Google Scholar] [CrossRef]
- Pavani, R.S.; Fernandes, C.; Perez, A.M.; Vasconcelos, E.J.R.; Siqueira-Neto, J.L.; Fontes, M.R.; Cano, M.I.N. RPA-1 from Leishmania amazonensis (LaRPA-1) Structurally Differs from Other Eukaryote RPA-1 and Interacts with Telomeric DNA via Its N-Terminal OB-Fold Domain. FEBS Lett. 2014, 588, 4740–4748. [Google Scholar] [CrossRef] [Green Version]
- Cuesta, J.; Read, M.A.; Neidle, S. The design of G-quadruplex ligands as telomerase inhibitors. Mini Rev. Med. Chem. 2003, 3, 11–21. [Google Scholar] [CrossRef]
- De Cian, A.; Grellier, P.; Mouray, E.; Depoix, D.; Bertrand, H.; Monchaud, D.; Teulade-Fichou, M.P.; Mergny, J.L.; Alberti, P. Plasmodium telomeric sequences: Structure, stability and quadruplex targeting by small compounds. Chembiochem 2008, 9, 2730–2739. [Google Scholar] [CrossRef]
- Morea, E.G.O.; Viviescas, M.A.; Fernandes, C.A.H.; Matioli, F.F.; Lira, C.B.B.; Fernandez, M.F.; Moraes, B.S.; da Silva, M.S.; Storti, C.B.; Fontes, M.R.M.; et al. A Calmodulin-like Protein (LCALA) Is a New Leishmania Amazonensis Candidate for Telomere End-Binding Protein. Biochim. Biophys. Acta BBA-Gen. Subj. 2017, 1861, 2583–2597. [Google Scholar] [CrossRef]
- Xu, Y. Chemistry in Human Telomere Biology: Structure, Function and Targeting of Telomere DNA/RNA. Chem. Soc. Rev. 2011, 40, 2719. [Google Scholar] [CrossRef] [PubMed]
- Morea, E.G.O.; Vasconcelos, E.J.R.; Alves, C.S.; Giorgio, S.; Myler, P.J.; Langoni, H.; Azzalin, C.M.; Cano, M.I.N. Exploring TERRA During Leishmania major Developmental Cycle and Continuous in vitro Passages. Int. J. Biol. Macromol. 2021, 174, 573–586. [Google Scholar] [CrossRef]
- van Leeuwen, F.; Taylor, M.C.; Mondragon, A.; Moreau, H.; Gibson, W.; Kieft, R.; Borst, P. β-D-Glucosyl-Hydroxymethyluracil Is a Conserved DNA Modification in Kinetoplastid Protozoans and Is Abundant in Their Telomeres. Proc. Natl. Acad. Sci. USA 1998, 95, 2366–2371. [Google Scholar] [CrossRef] [Green Version]
- Pays, E.; Delauw, M.F.; Laurent, M.; Steinert, M. Possible DNA Modification in GC Dinucleotides of Trypanosoma Brucei Telomeric Sequences; Relationship with Antigen Gene Transcriptiond. Nucleic Acids Res. 1984, 12, 5235–5247. [Google Scholar] [CrossRef] [PubMed]
- Bernards, A.; van Harten-Loosbroek, N.; Borst, P. Modification of Telomeric DNA in Trypanosoma Brucei; a Role in Antigenic Variation? Nucleic Acids Res. 1984, 12, 4153–4170. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwen, F.; Wijsman, E.R.; Kieft, R.; van der Marel, G.A.; van Boom, J.H.; Borst, P. Localization of the Modified Base J in Telomeric VSG Gene Expression Sites of Trypanosoma Brucei. Genes Dev. 1997, 11, 3232–3241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borst, P.; Sabatini, R. Base J: Discovery, Biosynthesis, and Possible Functions. Annu. Rev. Microbiol. 2008, 62, 235–251. [Google Scholar] [CrossRef]
- Shay, J.W.; Wright, W.E. Telomeres and Telomerase: Three Decades of Progress. Nat. Rev. Genet. 2019, 20, 299–309. [Google Scholar] [CrossRef]
- Weinrich, S.L.; Pruzan, R.; Ma, L.; Ouellette, M.; Tesmer, V.M.; Holt, S.E.; Bodnar, A.G.; Lichtsteiner, S.; Kim, N.W.; Trager, J.B.; et al. Reconstitution of Human Telomerase with the Template RNA Component HTR and the Catalytic Protein Subunit HTRT. Nat. Genet. 1997, 17, 498–502. [Google Scholar] [CrossRef]
- Beattie, T.L.; Zhou, W.; Robinson, M.O.; Harrington, L. Reconstitution of Human Telomerase Activity in Vitro. Curr. Biol. 1998, 8, 177–180. [Google Scholar] [CrossRef] [Green Version]
- Harrington, L. Biochemical Aspects of Telomerase Function. Cancer Lett. 2003, 194, 139–154. [Google Scholar] [CrossRef]
- Cano, M.I.N.; Dungan, J.M.; Agabian, N.; Blackburn, E.H. Telomerase in Kinetoplastid Parasitic Protozoa. Proc. Natl. Acad. Sci. USA 1999, 96, 3616–3621. [Google Scholar] [CrossRef] [Green Version]
- Giardini, M.A.; Fernández, M.F.; Lira, C.B.B.; Cano, M.I.N. Leishmania amazonensis: Partial Purification and Study of The Biochemical Properties of The Telomerase Reverse Transcriptase Activity from Promastigote-stage. Exp. Parasitol. 2011, 127, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Giardini, M.A.; Lira, C.B.B.; Conte, F.F.; Camillo, L.R.; de Siqueira Neto, J.L.; Ramos, C.H.I.; Cano, M.I.N. The Putative Telomerase Reverse Transcriptase Component of Leishmania Amazonensis: Gene Cloning and Characterization. Parasitol. Res. 2006, 98, 447–454. [Google Scholar] [CrossRef]
- Collins, K. Single-Stranded DNA Repeat Synthesis by Telomerase. Curr. Opin. Chem. Biol. 2011, 15, 643–648. [Google Scholar] [CrossRef] [Green Version]
- Rouda, S.; Skordalakes, E. Structure of the RNA-Binding Domain of Telomerase: Implications for RNA Recognition and Binding. Structure 2007, 15, 1403–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Peng, Y.; Mian, I.S.; Lue, N.F. Identification of Functionally Important Domains in the N-Terminal Region of Telomerase Reverse Transcriptase. Mol. Cell. Biol. 2000, 20, 5196–5207. [Google Scholar] [CrossRef] [Green Version]
- Armbruster, B.N.; Banik, S.S.R.; Guo, C.; Smith, A.C.; Counter, C.M. N-Terminal Domains of the Human Telomerase Catalytic Subunit Required for Enzyme Activity in vivo. Mol. Cell. Biol. 2001, 21, 7775–7786. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Brown, A.F.; Wu, J.; Xue, J.; Bley, C.J.; Rand, D.P.; Wu, L.; Zhang, R.; Chen, J.J.-L.; Lei, M. Structural Basis for Protein-RNA Recognition in Telomerase. Nat. Struct. Mol. Biol. 2014, 21, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Jansson, L.I.; Akiyama, B.M.; Ooms, A.; Lu, C.; Rubin, S.M.; Stone, M.D. Structural Basis of Template-Boundary Definition in Tetrahymena Telomerase. Nat. Struct. Mol. Biol. 2015, 22, 883–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Chan, H.; Cash, D.D.; Miracco, E.J.; Ogorzalek Loo, R.R.; Upton, H.E.; Cascio, D.; O’Brien Johnson, R.; Collins, K.; Loo, J.A.; et al. Structure of Tetrahymena Telomerase Reveals Previously Unknown Subunits, Functions, and Interactions. Science 2015, 350, aab4070. [Google Scholar] [CrossRef] [Green Version]
- Chan, H.; Wang, Y.; Feigon, J. Progress in Human and Tetrahymena Telomerase Structure Determination. Annu. Rev. Biophys. 2017, 46, 199–225. [Google Scholar] [CrossRef] [Green Version]
- Robart, A.R.; Collins, K. Human Telomerase Domain Interactions Capture DNA for TEN Domain-Dependent Processive Elongation. Mol. Cell 2011, 42, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Egan, E.D.; Collins, K. Biogenesis of Telomerase Ribonucleoproteins. RNA 2012, 18, 1747–1759. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, R.; Sanford, S.; Basu, S.; Park, M.; Pandya, U.M.; Li, B.; Chakrabarti, K. A Trans-Spliced Telomerase RNA Dictates Telomere Synthesis in Trypanosoma Brucei. Cell Res. 2013, 23, 537–551. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, J.C.; Cech, T.R. Human Telomerase: Biogenesis, Trafficking, Recruitment, and Activation. Genes Dev. 2015, 29, 1095–1105. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, K.T.T.T.; Wong, J.M.Y. Telomerase Biogenesis and Activities from the Perspective of Its Direct Interacting Partners. Cancers 2020, 12, 1679. [Google Scholar] [CrossRef]
- Viviescas, M.A.; Cano, M.I.N.; Segatto, M. Chaperones and Their Role in Telomerase Ribonucleoprotein Biogenesis and Telomere Maintenance. Curr. Proteom. 2018, 16, 31–43. [Google Scholar] [CrossRef]
- Weber, H.; Valbuena, J.R.; Barbhuiya, M.A.; Stein, S.; Kunkel, H.; García, P.; Bizama, C.; Riquelme, I.; Espinoza, J.A.; Kurtz, S.E.; et al. Small Molecule Inhibitor Screening Identified HSP90 Inhibitor 17-AAG as Potential Therapeutic Agent for Gallbladder Cancer. Oncotarget 2017, 8, 26169–26184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nepomuceno-Mejía, T.; Florencio-Martínez, L.E.; Martinez-Calvillo, S. Nucleolar Division in the Promastigote Stage of Leishmania major Parasite: A Nop56 Point of View. Biomed. Res. Int. 2018, 2018, 1641839. [Google Scholar] [CrossRef] [PubMed]
- Zamudio, J.R.; Mittra, B.; Chattopadhyay, A.; Wohlschlegel, J.A.; Sturm, N.R.; Campbell, D.A. Trypanosoma brucei Spliced Leader RNA Maturation by the Cap 1 2′-O-Ribose Methyltransferase and SLA1 H/ACA SnoRNA Pseudouridine Synthase Complex. Mol. Cell. Biol. 2009, 29, 1202–1211. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Kolet, L.; Doniger, T.; Biswas, V.K.; Unger, R.; Tzfati, Y.; Michaeli, S. The Trypanosoma brucei Telomerase RNA (TER) Homologue Binds Core Proteins of the C/D SnoRNA Family. FEBS Lett. 2013, 587, 1399–1404. [Google Scholar] [CrossRef] [Green Version]
- Greider, C. Telomerase Is Processive. Mol. Cell. Biol. 1991, 11, 4572–4580. [Google Scholar] [CrossRef]
- Xin, H.; Liu, D.; Wan, M.; Safari, A.; Kim, H.; Sun, W.; O’Connor, M.; Songyang, Z. TPP1 Is a Homologue of Ciliate TEBP-beta And Interacts with POT1 to Recruit Telomerase. Nature 2007, 445, 559–562. [Google Scholar] [CrossRef]
- Latrick, C.M.; Cech, T.R. POT1–TPP1 Enhances Telomerase Processivity by Slowing Primer Dissociation and Aiding Translocation. EMBO J. 2010, 29, 924–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aramburu, T.; Plucinsky, S.; Skordalakes, E. POT1-TPP1 Telomere Length Regulation and Disease. Comput. Struct. Biotechnol. J. 2020, 18, 1939–1946. [Google Scholar] [CrossRef]
- Chiodi, I.; Mondello, C. Telomere-Independent Functions of Telomerase in Nuclei, Cytoplasm, and Mitochondria. Front. Oncol. 2012, 2, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campelo, R.; Díaz Lozano, I.; Figarella, K.; Osuna, A.; Ramírez, J.L. Leishmania Major Telomerase TERT Protein Has a Nuclear/Mitochondrial Eclipsed Distribution That Is Affected by Oxidative Stress. Infect. Immun. 2015, 83, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagoubat, A.; Corrales, R.M.; Bastien, P.; Lévêque, M.F.; Sterkers, Y. Gene Editing in Trypanosomatids: Tips and Tricks in the CRISPR-Cas9 Era. Trends Parasitol. 2020, 36, 745–760. [Google Scholar] [CrossRef] [PubMed]
- Yagoubat, A.; Crobu, L.; Berry, L.; Kuk, N.; Lefebvre, M.; Sarrazin, A.; Bastien, P.; Sterkers, Y. Universal Highly Efficient Conditional Knockout System in Leishmania, with a Focus on Untranscribed Region Preservation. Cell. Microbiol. 2020, 22, e13159. [Google Scholar] [CrossRef]
- Beneke, T.; Gluenz, E. LeishGEdit: A Method for Rapid Gene Knockout and Tagging Using CRISPR-Cas9. In Leishmania; Clos, J., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 1971, pp. 189–210. ISBN 978-1-4939-9209-6. [Google Scholar]
- Xi, L.; Schmidt, J.C.; Zaug, A.J.; Ascarrunz, D.R.; Cech, T.R. A Novel Two-step Genome Editing Strategy with CRISPR-Cas9 Provides New Insights into Telomerase Action and TERT Gene Expression. Genome Biol. 2015, 16, 231. [Google Scholar] [CrossRef] [Green Version]
- Xia, B.; Amador, G.; Viswanatha, R.; Zirin, J.; Mohr, S.E.; Perrimon, N. CRISPR-Based Engineering of Gene Knockout Cells by Homology-Directed Insertion in Polyploid Drosophila S2R+ Cells. Nat. Protoc. 2020, 15, 3478–3498. [Google Scholar] [CrossRef] [PubMed]
- Dreesen, O.; Li, B.; Cross, G. Telomere Structure and Shortening in Telomerase-deficient Trypanosoma brucei. Nucleic Acids Res. 2005, 33, 4536–4543. [Google Scholar] [CrossRef] [PubMed]
- Haendeler, J.; Hoffmann, J.; Rahman, S.; Zeiher, A.M.; Dimmeler, S. Regulation of Telomerase Activity and Anti-Apoptotic Function by Protein-Protein Interaction and Phosphorylation. FEBS Lett. 2003, 536, 180–186. [Google Scholar] [CrossRef]
- Lee, J.; Sung, Y.H.; Cheong, C.; Choi, Y.S.; Jeon, H.K.; Sun, W.; Hahn, W.C.; Ishikawa, F.; Lee, H.-W. TERT Promotes Cellular and Organismal Survival Independently of Telomerase Activity. Oncogene 2008, 27, 3754–3760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Zheng, C.; Hou, M.; Lindvall, C.; Li, K.-J.; Erlandsson, F.; Björkholm, M.; Gruber, A.; Blennow, E.; Xu, D. Deletion of the Telomerase Reverse Transcriptase Gene and Haploinsufficiency of Telomere Maintenance in Cri Du Chat Syndrome. Am. J. Hum. Genet. 2003, 72, 940–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Ding, D.; Wang, M.; Cong, Y.-S. Telomerase Reverse Transcriptase in the Regulation of Gene Expression. BMB Rep. 2014, 47, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Liu, W.; Chen, C.; Zhang, H.; Yue, D.; Li, C.; Zhang, L.; Gao, L.; Huo, Y.; Liu, C.; et al. TPP1 OB-Fold Domain Protein Suppresses Cell Proliferation and Induces Cell Apoptosis by Inhibiting Telomerase Recruitment to Telomeres in Human Lung Cancer Cells. J. Cancer Res. Clin. Oncol. 2019, 145, 1509–1519. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assis, L.H.C.; Andrade-Silva, D.; Shiburah, M.E.; de Oliveira, B.C.D.; Paiva, S.C.; Abuchery, B.E.; Ferri, Y.G.; Fontes, V.S.; de Oliveira, L.S.; da Silva, M.S.; et al. Cell Cycle, Telomeres, and Telomerase in Leishmania spp.: What Do We Know So Far? Cells 2021, 10, 3195. https://doi.org/10.3390/cells10113195
Assis LHC, Andrade-Silva D, Shiburah ME, de Oliveira BCD, Paiva SC, Abuchery BE, Ferri YG, Fontes VS, de Oliveira LS, da Silva MS, et al. Cell Cycle, Telomeres, and Telomerase in Leishmania spp.: What Do We Know So Far? Cells. 2021; 10(11):3195. https://doi.org/10.3390/cells10113195
Chicago/Turabian StyleAssis, Luiz H. C., Débora Andrade-Silva, Mark E. Shiburah, Beatriz C. D. de Oliveira, Stephany C. Paiva, Bryan E. Abuchery, Yete G. Ferri, Veronica S. Fontes, Leilane S. de Oliveira, Marcelo S. da Silva, and et al. 2021. "Cell Cycle, Telomeres, and Telomerase in Leishmania spp.: What Do We Know So Far?" Cells 10, no. 11: 3195. https://doi.org/10.3390/cells10113195